|

- 现金
- 62111 元
- 精华
- 26
- 帖子
- 30441
- 注册时间
- 2009-10-5
- 最后登录
- 2022-12-28

|
4.1.1. Serum Empty Virions (HBcAg) as a Marker for Hepatic CCC DNA
Since the production of empty virions is uncoupled from viral DNA replication but requires the expression of both HBcAg and HBsAg, they can in principle serve as an effective biomarker for transcriptionally active CCC DNA during current antiviral therapy with RT inhibitors, which, as mentioned above, can reduce serum HBV DNA (i.e., complete virions) to undetectable levels but has no direct effect on CCC DNA level or its transcriptional activity [78,80]. As mentioned above, a minority of the treated patients do experience significant decreases in hepatic CCC DNA with current antiviral therapy, presumably due to host-mediated elimination of infected
cells or CCC DNA and the therapy-induced elimination of the CCC DNA precursor (i.e., RC DNA) and consequently, diminished replenishment of the CCC DNA pool [78,81,82,83]. In a pilot study to evaluate the potential of serum empty virions as a surrogate biomarker for hepatic CCC DNA, we determined serum levels of empty virions, using a relatively insensitive western blot assay (with a detection limit of ca. 50 ng/mL), together with viral DNA (complete virions) and HBsAg levels, in a small group of patients who underwent treatment with the RT inhibitor tenofovir [9]. Levels of serum empty virions can be monitored conveniently by measuring serum HBcAg levels, as the amount of HBcAg in complete virions contributes to less than 1% of the total serum HBcAg (i.e., from both complete and empty virions).
Before tenofovir treatment, serum empty virions were present at levels up to 1011/mL and at more than 50- to 100,000-fold excess compared to RC DNA-containing virions [9], consistent with our earlier observations in HBV-infected chimpanzees and in the supernatants of HBV-replicating hepatoma cells [8]. Following the antiviral therapy, the secretion of complete virions decreased dramatically (by ca. 107/mL or more) in virtually all patients, as expected. However, secretion of empty virions was not decreased in most cases even after years of potent HBV DNA suppression. This is, of course, exactly as predicted given that DNA synthesis is required for secretion of complete virions, but completely dispensable for empty virion secretion, which would continue unabated unless the hepatic CCC DNA is eliminated or stably silenced. Significant reductions in, and possibly complete loss of, serum empty virions were in fact observed in a minority of treated patients, including those who achieved HBsAg loss. The results suggest that these patients might indeed have experienced significant reductions or loss in hepatic CCC DNA levels (or CCC DNA transcriptional activity) after the therapy, although the unavailability of the corresponding liver tissues unfortunately precluded a direct measurement of hepatic CCC DNA in these patients. Future studies with larger sample sizes and more sensitive HBcAg assay format, together with direct measurements of corresponding hepatic CCC DNA levels, will be needed to further assess the usefulness of serum empty virions as a surrogate marker for hepatic CCC DNA and to determine the potential diagnostic and prognostic significance of serum empty virions. As the HBc sequence is known to be more variable in some regions than in the others [84], epitopes will have to be carefully considered for antibody-based assays to detect serum HBcAg (i.e., empty virions).
Interestingly, attempts were made over two-decade ago to measure serum HBc, before the realization of empty virion secretion. An enzyme-linked immunosorbent assay (ELISA) kit was developed to measure serum HBcAg, using an antibody specific for the HBc CTD, which indeed showed a good correlation between serum HBc and HBV DNA in untreated patients, and furthermore, the decrease of serum HBc was much less than that of serum HBV DNA during treatment with an RT inhibitor (lamivudine in this case) in the single patient who was monitored [85]. These results led the authors to speculate the secretion of DNA-free virions although they didn't provide any other supporting evidence. In the meantime, a different ELISA kit was developed to measure the so-called HBcrAg in serum samples, which has been used in a number of clinical studies as a putative surrogate in attempt to monitor hepatic CCC DNA.
It is important to point out that some confusions exist in the literature as to what “HBcrAg” is exactly. Initially, in 2002, an ELISA kit was reported for the detection of both the soluble HBeAg and HBcAg that was released from virions in serum samples, using antibodies targeted to the sequences shared by both HBc and HBe [86]. A few years later, an aberrant HBeAg and HBcAg related protein, the so-called p22cr (with an even longer N-terminal extension than HBe and missing the CTD of HBc; see above), was claimed to be present in DNA-free HBV virions in serum samples from HBV infected individuals. This aberrant protein was also named HBcrAg. In the literature, the term “HBcrAg” has been used to describe a combination of HBc and HBe [13,86,87,88]; the supposedly aberrant p22cr [47]; and a combination of all three entities, HBc, HBe, and p22cr [89,90,91]. As discussed above, the so-called p22cr probably does not really exist and the current HBcrAg ELISA kit most likely detects a combination of HBcAg (released from virions) and HBeAg. Thus, with HBeAg present, the HBcrAg kit detects both serum empty virions and HBeAg but in the absence of HBeAg, it essentially detects empty virions (again with the contribution of serum HBcAg from complete virions being negligible).
Not surprisingly, serum HBcrAg levels were found to correlate relatively well with serum HBV DNA and HBsAg levels, to decrease in HBeAg (−) phase, and to decrease much slower than serum HBV DNA and remains detectable in serum HBV DNA (−) patients upon nucleoside reverse transcriptase inhibitor (NRTI) treatment [13,88,90], as we observed for serum empty virions [9]. Furthermore, serum HBcrAg levels were correlated with CCC DNA levels and frequency of HBcAg (+) hepatocytes in the liver and with liver disease activity [87,89,92,93]. It was also found to predict liver cancer development and recurrence in HBV-infected patients, and was an even better predictor than serum HBV DNA [92,94]. Serum HBcrAg was also found to associate with reactivation of occult hepatitis B following immunosuppressive therapy [91].
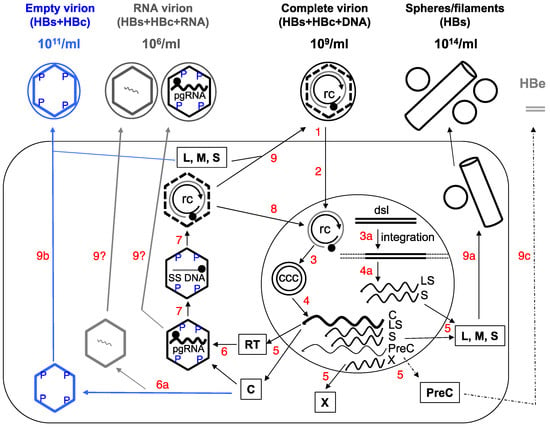
As discussed above, serum HBeAg levels may not strictly correlate with hepatic CCC DNA levels or its transcriptional activity, due to the frequent mutations in the PreC region and core promoter. Therefore, measurement of both serum HBeAg and HBcAg, by using the HBcrAg assay, may not entirely reflect functional CCC DNA in the liver, since CCC DNA defective in HBeAg expression can nevertheless still express HBcAg and the other viral proteins and sustain HBV persistence. Given the aberrant “p22cr” or “HBcrAg” is most likely an artifact, as discussed above, it will be advantageous to measure exclusively serum HBcAg (i.e., empty virions), without the variable contribution of HBeAg. Moreover, as mentioned above, the ratio of empty vs. complete virions in different patients varies widely. Among other factors, this ratio may reflect the efficiency of intrahepatic assembly of empty vs. pgRNA-containing capsids, which, together with the efficiency of reverse transcription and virion assembly and secretion, ultimately determine the ratio of empty vs. complete virions in the blood (Figure 1). Thus, measurement of both complete and empty virions in the blood can also help monitor these events in the liver, which could reflect the changing viral activities, host physiology, and virus-host interactions.
4.1.1。血清空病毒(HBcAg)作为肝癌CCC DNA的标记物
由于空病毒粒子的产生与病毒DNA复制物分离,而是需要HBcAg和HBsAg两者的表达,它们原则上可以作为转录活性CCC DNA的有效生物标志物,在目前用RT抑制剂的抗病毒治疗中,如上所述,可以将血清HBV DNA(即完整病毒体)降低到不可检测的水平,但对CCC DNA水平或其转录活性没有直接影响[78,80]。如上所述,目前的抗病毒治疗中,少数治疗的患者确实经历肝CCC DNA的显着降低,这可能是由于宿主介导的感染细胞或CCC DNA的消除以及治疗诱导的CCC DNA前体的消除(即, RC DNA),从而减少CCC DNA池的补充[78,81,82,83]。在一项试验性研究中,评估血清空病毒颗粒作为肝癌CCC DNA替代生物标志物的潜力,我们使用相对不敏感的蛋白质印迹法(检测限为约50ng / mL)测定空白病毒粒子的血清水平,连同病毒DNA(完整病毒体)和HBsAg水平,在一小组接受RT抑制剂替诺福韦治疗的患者中[9]。通过测量血清HBcAg水平可以方便地监测血清空病毒体的水平,因为完整病毒粒子中HBcAg的量有助于小于总血清HBcAg的1%(即来自完整的和空的病毒粒子)。
在替诺福韦治疗前,血清空白病毒以高达1011 / mL的水平存在,与含RC DNA的病毒体相比超过50至100,000倍[9],这与我们以前在HBV感染黑猩猩和HBV复制肝癌细胞的上清液[8]。在抗病毒治疗之后,如预期的那样,完全病毒体的分泌在几乎所有患者中显着降低(约107 / mL或更多)。然而,即使经过多年的有效的HBV DNA抑制,空肠病毒的分泌在大多数情况下也没有减少。这当然是完全一样预测的,因为DNA合成是分泌完整病毒粒子所必需的,但对于空的病毒粒子分泌是完全无用的,除非肝CCC DNA被消除或稳定沉默,否则它将继续有增无减。事实上,在少数治疗患者中观察到血清空病毒体的显着减少,甚至可能完全丧失,包括那些获得HBsAg损失的患者。结果表明,这些患者可能确实在治疗后肝脏CCC DNA水平(或CCC DNA转录活性)显着减少或减少,尽管相应的肝组织的不可用性不幸地排除了这些患者肝脏CCC DNA的直接测量。将需要具有较大样本量和更敏感的HBcAg测定形式的未来研究以及相应的肝CCC DNA水平的直接测量,以进一步评估血清空病毒粒子作为肝CCC DNA的替代标志物的有用性并确定潜在诊断和血清空病毒粒子的预后意义。由于已知HBc序列在某些区域比其他区域更为可变[84],因此需要认真考虑表位以进行基于抗体的检测以检测血清HBcAg(即空的病毒粒子)。
有趣的是,在实现空白病毒粒子分泌之前,在二十年前尝试测量血清HBc。开发了一种酶联免疫吸附测定(ELISA)试剂盒,用HBc CTD特异性抗体测定血清HBcAg,在未经治疗的患者中,血清HBc和HBV DNA的相关性确实显示出良好的相关性,此外,血清HBc在被监测的单个患者中使用RT抑制剂(在这种情况下为拉米夫定)治疗期间,血清HBV DNA的含量远低于[85]。这些结果导致作者推测无DNA病毒粒子的分泌,尽管它们没有提供任何其他证据。同时,开发了一种不同的ELISA试剂盒,用于测量血清样品中所谓的HBcrAg,已被用于许多临床研究,作为试图监测肝脏CCC DNA的推定替代物。
重要的是要指出,文献中存在一些关于“HBcrAg”是什么的混乱。最初,在2002年,报告了ELISA试剂盒,用于检测从血清样品中的病毒体释放的可溶性HBeAg和HBcAg,使用针对HBc和HBe共有序列的抗体[86]。几年之后,所谓的无异常HBeAg和HBcAg相关蛋白被称为存在于无DNA的HBV中,所谓的p22cr(具有比HBe更长的N-末端延伸和缺失HBc的CTD,见上文)来自HBV感染个体的血清样品中的病毒粒子。这种异常蛋白质也被命名为HBcrAg。在文献中,术语“HBcrAg”用于描述HBc和HBe的组合[13,86,87,88];据说是异常的p22cr [47];和所有三个实体的组合,HBc,HBe和p22cr [89,90,91]。如上所述,所谓的p22cr可能不存在,目前的HBcrAg ELISA试剂盒最有可能检测到HBcAg(从病毒体释放)和HBeAg的组合。因此,当HBeAg存在时,HBcrAg试剂盒检测血清空白病毒粒子和HBeAg,但是在没有HBeAg的情况下,它基本上可以检测到空的病毒粒子(再次,来自完整病毒粒子的血清HBcAg的贡献可以忽略不计)。
不出人意料的是,发现血清HBcrAg水平与血清HBV DNA和HBsAg水平相关性相对较低,HBeAg( - )期降低,并且比血清HBV DNA慢得多,并且在血清HBV DNA( - ))患者中仍然可检测到核苷逆转录酶抑制剂(NRTI)治疗[13,88,90],我们观察到血清空病毒体[9]。此外,血清HBcrAg水平与肝脏中HBcAg(+)肝细胞的CCC DNA水平和频率以及肝脏疾病活动相关[87,89,92,93]。还发现预测HBV感染患者的肝癌发展和复发,是比血清HBV DNA更好的预测指标[92,94]。还发现血清HBcrAg与免疫抑制治疗后隐匿性乙型肝炎的再激活相关[91]。
如上所述,由于PreC区域和核心启动子的频繁突变,血清HBeAg水平可能不与肝CCC DNA水平或其转录活性严格相关。因此,通过使用HBcrAg测定,血清HBeAg和HBcAg两者的测量可能不完全反映肝脏中的功能性CCC DNA,因为HBeAg表达中的CCC DNA缺陷仍然可以表达HBcAg和其他病毒蛋白并维持HBV持久性。鉴于异常的“p22cr”或“HBcrAg”最有可能是上述讨论的神器,仅测量血清HBcAg(即空的病毒粒子)是有利的,而HBeAg的贡献不大。此外,如上所述,不同患者的空白对完全病毒粒子的比例差别很大。除了其他因素之外,该比例可能反映了空载体与含有pgRNA的衣壳的肝内装配效率,其与反转录和病毒粒子装配和分泌的效率一起最终决定了血液与空白对完全病毒粒子的比例(图1)。因此,血液中完整和空的病毒粒子的测量也可以帮助监测肝脏中的这些事件,这可以反映病毒活动的变化,宿主生理学和病毒 - 宿主相互作用。 |
|



 发表于 2017-3-29 20:54
发表于 2017-3-29 20:54