1Clinical Research Directorate/Clinical Monitoring Research Program, Leidos Biomedical Research Inc., (formerly SAIC-Frederick, Inc.) Frederick National Laboratory for Cancer Research, Frederick, MD 21702, USA
2Laboratory of Immunoregulation, National Institute of Allergy & Infectious Diseases, NIH, Department of Health & Human Services, Bethesda, MD 20892, USA
Approximately 240 million people worldwide have chronic HBV (CHB).[1] Current therapy for HBV is aimed at achieving suppression of HBV replication at levels below detection. Although this can be accomplished in almost all patients, long-term management of CHB remains a challenge. Failure to achieve sustained response and HBV persistence is related to the viral factors and inadequate induction of immune response that are seen in acute HBV patients, which naturally clears the infection. HLA polymorphisms also determine the variability in host immune response by influencing host susceptibility to HBV infection.
Advancement in the understanding of the basis of HBV persistence has guided the development of strategies that could lead to a functional cure for HBV infection. We discuss the potential strategies under development to achieve a functional cure of hepatitis B by targeting the virus, host or both. Some of these interventions are currently experimental and some have attained preclinical validation, whereas few have reached active clinical trials at this time.
Pitfalls of Current Therapy Chronic Suppression Without Sustained Cure
Current antiviral therapies are aimed towards inhibition of viral replication. Sustained viral suppression is associated with improved outcomes and it has been shown that elevated HBV DNA level is a strong predictor of increased risk of cirrhosis[13] and HCC[14,15] in CHB. None of the current therapies lead to HBsAg loss or seroconversion in the majority of patients. In addition, current antiviral therapies do not target defective immune response and persistence of covalently closed circular DNA (cccDNA) in the infected hepatocytes. Thus, the goal of current treatment is to achieve long-term virologic control since elimination or 'cure' is not possible.
Despite the tremendous improvement in CHB therapy with new antivirals, viral replication typically rebounds after the treatment is stopped. Therefore, complete elimination or cure is still not possible with current treatment. The plausible explanation is the persistence of cccDNA, which plays a vital role in persistence and reactivation. Nucleoside analog therapy prevents further formation of cccDNA, but has no effect on existing cccDNA. Ideally, elimination of HBV can be defined by loss of HBsAg and seroconversion to anti-HBs antibody and sustained suppression of HBV DNA. This may also result in depletion or inactivation of cccDNA. Until we reach the point when elimination/eradication of cccDNA can be achieved, HBsAg can be used as a marker of surrogate for the level of transcriptionally active cccDNA. Several studies have shown a positive correlation between transcriptionally active cccDNA and HBsAg levels in CHB patients.[24,25]
Understanding the basis for HBV persistence is critical in designing therapeutic strategies to eradicate HBV. Chronic HBV infection is characterized by an evolving interplay between viral replication and host immune responses. Both viral and host factors contribute to persistence of HBV. Highly efficient and unique replication mechanism of the virus uses a transcriptional template, cccDNA that is sequestered inside the nucleus, and escapes detection by innate DNA sensing cellular machinery. Another factor is the production of viral proteins (HBsAg, HBeAg), which function as a tolerogen and leads to T-cell exhaustion.[26] Advancement in the understanding of the mechanisms involved in HBV persistence has helped to develop strategies that overcome these factors and could result in sustained virologic remission. 乙肝病毒持续性的基础
Error Prone Replication. HBV is a small, enveloped DNA virus with a very unique genomic organization and replication mechanism. Genome length of HBV is only 3200 bp compared with 10,000 bp for HIV and that multiple overlapping open reading frames (ORFs) may impose more constraints against variation on HBV than HIV. Despite the constraint imposed by ORFs, HBV replication is error prone due to lack of proofreading activity of HBV polymerase (error rate of 10−4 to 10−5), which leads to an accumulation of a pool of genomic sequences with heterogeneous viral population, also called quasispecies.[17,27] These viral variants have a robust survival advantage in particular, when exposed to multiple selection pressures such as immunological pressure from hepatitis B immunoglobulin, NAs and/or vaccination. 容易出错的复制。乙肝病毒是一种体积小,包膜DNA病毒,具有非常独特的基因组结构和复制的机制。 10,000 bp的艾滋病毒和多个重叠的开放阅读框(ORF),可并处更多的约束对变异的乙肝病毒比乙肝病毒相比,基因组的长度只有3200个基点。尽管通过的ORF所施加的约束,HBV复制是容易出错的,由于缺乏HBV聚合酶的校对活性的(10-4〜10-5的误码率),这导致了与异构病毒种群的基因组序列的池的积累,也称为准种[17,27],这些病毒变异体具有坚固的生存优势,特别是当暴露于多个选择压力,例如从B型肝炎免疫球蛋白,NAS和/或疫苗接种的免疫压力。
Precore/Core Mutants. The viral core mRNA encodes a core protein (major nucleocapsid protein), DNA polymerase (which reverse transcribes RNA pregenome), and serves as pregenomic RNA, which acts as a template for reverse transcription. Precore mRNA encodes the precore protein, which is processed in the endoplasmic reticulum (ER) to produce HBeAg; the basal core promoter (BCP), nucleotide 1744–1804, resides in X ORF, and controls transcription of both precore and core regions.[28] A variety of precore and core mutants have been reported. There are two well-studied precore mutations: stop codon mutation at nt 1896, which results in cessation of HBeAg expression, and a mutation in BCP at nt 1762 and nt 1764, which results in diminished production of HBeAg and a resulting increased host immune response.[28] These mutations lead to the development of HBeAg-negative CHB. Associations of precore mutants and increased pathogenicity have been described. Earlier studies have demonstrated that precore mutants might be associated with severe chronic liver disease and with acute liver failure. It has been shown that patients with detectable precore and/or BCP mutants have a lower probability of response and are less optimal candidates for PEG-IFN therapy.[29] Double mutations in BCP at nt 1762 and nt 1764 are reported to be associated with severe liver disease,[30] fulminant hepatitis,[31] cirrhosis and HCC.[32,33] 前C区/核心突变体。病毒核心基因编码核心蛋白(主要衣壳蛋白),DNA聚合酶(其中反向转录的RNA前基因组),并作为前基因组RNA,其作为用于反转录的模板。前C区基因编码的前核心蛋白,其在内质网(ER),以产生HBeAg的处理;基底核心启动子(BCP),核苷酸1744年至1804年,位于X中的ORF,并控制这两个前C区和芯区的转录。[28]各种前C区和核心突变体已有报道。有两个充分研究前C区突变:终止密码突变新台币1896年,这将导致停止HBeAg的表达,而在BCP突变,至新台币1762 NT1764年,这将导致生产减少的e抗原和由此增加的宿主免疫反应[28]这些突变导致HBeAg阴性慢性乙型肝炎的发展。前C区突变体,增加致病的关联已被描述。早先的研究已经表明,前C区突变体可能具有严重的慢性肝脏疾病和急性肝衰竭有关。它已经表明,患者检测前C区和/或BCP变异有反应的机率较低,而且不太理想人选PEG-IFN治疗。[29]双突变的BCP,至新台币1762和NT1764顷报道与相关严重的肝脏疾病,[30]暴发性肝炎,[31]肝硬化和肝癌。[32,33]
HBV Genotypes. HBV genotypes account for the heterogeneity in clinical manifestations and treatment response among patients with chronic hepatitis B in different parts of the world; several studies reported correlation of HBV genotype with clinical outcomes and response to treatment, especially IFN treatment.[34] To date, ten HBV genotypes (A–J) and several subtypes have been identified, defined by divergence in the entire HBV genomic sequences and distinct geographic distribution. Genotype A is found as an independent risk factor for progression to chronic infection and persistence following acute hepatitis B infection.[35] Acute infection with genotypes A and D results in higher rates of chronicity than genotypes B and C. Patients with genotypes C and D have lower rates of spontaneous HBeAg seroconversion as compared with genotype A and B. There is also a clear association between HBV genotypes, and precore and BCP mutations. Genotype C has a higher frequency of double mutation in BCP A1762T/G1764A, pre-S deletion and is associated with higher viral load than genotype B. Similarly, genotype D has a higher prevalence of BCP A1762T/G1764A mutation than genotype A. Genotype C and D are associated with more severe liver disease, including cirrhosis and HCC. Genotype A and B shows better responses to IFN-based therapy than genotypes C and D, but there are few consistent differences for NAs.[36] HBV基因型。 HBV基因型占在临床表现和患者中,在世界不同地区慢性乙型肝炎治疗反应的异质性;几项研究HBV基因型与临床结果和对治疗的反应,特别是IFN治疗的报道的相关性。[34]迄今,10 HBV基因型(A-J)和几种亚型已经确定,由发散在整个HBV基因组序列的定义和独特的地理分布。 A基因型发现,作为发展为慢性感染和持续性急性乙型肝炎感染的独立危险因素[35]。急性感染基因型A和D会导致慢性化率高于基因型B和C患者基因型C和D与A基因型比较,二也有HBV基因型之间有明显的联系,和前C区和BCP变异有自发性HBeAg血清学转换率较低。 C基因型在BCP A1762T/ G1764A,前S缺失双突变频率较高,并具有较高的病毒载量高于基因型B.同样相关,基因型D是BCP A1762T/ G1764A突变的发病率较高,比基因型A. C基因型和D是更严重的肝脏疾病,包括肝硬化和肝癌有关。基因型A和B具有更好的响应干扰素为基础的治疗比基因型C和D,但也有NAS的一些一致的差异。[36]
cccDNA. A major determinant in the slow kinetics of HBV clearance from infected cells and persistence is the presence of cccDNA. The HBV genome assumes a supercoiled configuration of cccDNA and exists in association with histones and DNA chaperone proteins as a minichromosome. This form allows HBV to persist inside the nucleus by avoiding host innate immune responses. Furthermore, infected hepatocytes have a long half-life, which allows the maintenance of cccDNA in the nuclei of infected cells indefinitely[37] and acts as reservoir for reactivation of viral genome replication. Studies have shown that drug-resistant mutations are archived in the cccDNA and can be rapidly selected out with the use of drugs that exhibits crossresistance.[38–40] The estimated 15–50 copies/cell of cccDNA in the nucleus serve as a store of viral escape variants generated by the error-prone viral polymerase.[41] Antiviral therapies with the currently approved antiviral agents suppress viral replication but do not directly target cccDNA. Thus, inactivation or elimination of cccDNA is one of the potential novel strategies for eradication of HBV. cccDNA的。在清除乙肝病毒,从病毒感染的细胞和持久性慢反应动力学的主要决定因素是cccDNA的存在。 HBV基因组假设的cccDNA的超螺旋结构和存在于与组蛋白和DNA的分子伴侣蛋白的微染色体的关联。这种形式可以让乙肝病毒,避免宿主天然免疫应答坚持的细胞核内。此外,感染的肝细胞具有较长的半衰期,这允许cccDNA的在感染细胞的细胞核的保养无限期[37],并作为贮存于病毒基因组复制的再活化。研究表明,药物抗性突变被存档在cccDNA的,并且可以迅速地筛选出与使用的药物表现出交叉耐受[38-40]估计15-50个拷贝/ cccDNA的在核细胞用作存储病毒逃避变体由易错的病毒聚合酶产生的。[41]抗病毒疗法与目前批准的抗病毒药物抑制病毒的复制,但不直接针对的cccDNA。因此,灭活的cccDNA或消除是消除HBV的潜在的新的策略之一。
作者: StephenW 时间: 2014-9-7 20:49
Host Factors宿主因素
Host Genetics. Persistence of HBV and variability in disease outcome also depends on the multiple host factors. HLA type of an individual is an important factor that determines the variability in host immune response to HBV. Evidence from genome-wide association studies has shown that the HLA DRB locus alleles DRref-1*1301/2 are consistently associated with spontaneous resolution of infection,[42] whereas HLA-DR7 (DRref-1*07) and HLA-DR3 (DRref-1*0301) were found to be associated with increased susceptibility to chronic HBV.[43–47] Other HLA types associated with risk of chronic infection were HLA-DPA1(*)0202-DPref-1(*)0501 and HLA-DPA1(*)0202-DPref-1(*)0301.[48] HLA-DRref-1*0701 and DRref-1*0301 have not only been associated with increased susceptibility to chronic infection but also with failure to respond to HBsAg-based vaccine.[49,50] Other alleles strongly associated with no/poor response are DRref-1*03, DRref-1*07, DQref-1*02 and DPref-1*1101.[51] HLA- DRref-1*0901, DQA1*0301, DQA1*0501 and DQref-1*0301 are found to be consistently associated with persistent HBV infection in different ethnic groups[52–56] Conceivably, this genotype influences host response by allowing more promiscuous binding of peptides to this allele than others, resulting in a broader T-cell response in subjects with the favorable allele and hence self-limiting infection.[57] 宿主遗传。乙肝病毒的持久性和变异性疾病的结果还取决于多种宿主因素。
个体
的HLA型,是决定在对HBV的宿主免疫反应的变异性的重要因素。从全基因组关联研究的证据表明,HLA DRB等位基因DRref-1 *二分之一千三百零一始终与感染的自发分辨率有关,[42]而HLA-DR7(DRref-1 *07)和HLA-DR3( DRref-1 *0301)被发现与增加的易感性的慢性HBV相关[43-47]对慢性感染的风险相关联的其他HLA型为HLA-DPA1(*)0202-DPref-1(*)0501和HLA -DPA1(*)0202-DPref-1(*),0301[48] HLA-DRref-1 *0701和DRref-1 *0301不仅具有增加的易感性的慢性感染被关联而且还与未能对HBsAg回应为基础的疫苗。无/低反应密切相关[49,50]的其他等位基因是DRref-1*03,DRref-1*07,DQref-1*02和DPref-1* 1101。[51] HLA-DRref- 1* 0901,DQA1 *0301,DQA1 *0501和DQref-1*0301顷发现是始终与HBV持续感染有关不同族群[52-56]可以想象,这种基因型的影响,允许肽更加淫乱的结合宿主反应该等位基因比其他,导致在受试者的有利等位基因,因而自限感染更广的T细胞应答。[57]
Another genome-wide linkage study in siblings with CHB found that a region of linkage on chromosome 21 with single nucleotide polymorphisms spanning the IFN-α receptor II and IL-10 receptor II was associated with chronicity.[57,58]
在兄弟姐妹慢性乙型肝炎的另一个全基因组关联研究发现,联动与跨越IFN-α受体II和IL-10受体II的单核苷酸多态性21号染色体的一个区域与慢性关联[57,58]
Host–Viral Interactions. Adaptive immune responses to HBV are blunted in CHB subjects when compared with those who have resolved acute infection. Studies have demonstrated that T cells responding to HBV antigens from these subjects have an exhausted phenotype and are less responsive to HBV antigens.[59] Furthermore, HBV antigens have been shown to interfere with innate immune recognition, by specifically hindering signaling of Toll-like receptor (TLR) 2, 7 and 9 molecules. These molecules are vital in generating an effective innate adaptive crosstalk that is the cornerstone of an effective anti-HBV immunity. 主机病毒的相互作用。当那些谁已经解决急性感染相比,适应性免疫应答对HBV有减弱慢性乙型肝炎科目。研究已经表明,T细胞从这些受试者响应HBV抗原有一个耗尽型,而且不太响应于乙型肝炎病毒的抗原。[59]此外,HBV抗原已被证明是干扰先天免疫识别,通过特异性阻碍的信令Toll样受体(TLR)2,7和9的分子。这些分子产生有效的先天性自适应串扰是一种有效的抗HBV免疫的基础是至关重要的。
只为你 Table 1. Viral factors. Factors Targets Results Ref. Viral entry (NTCP receptor at basolateral membrane of hepatocytes) Myrcludex-B Specifically targets NTCP and efficiently block infection
Phase II trial on CHB patients recently completed – awaiting results Cell culture: [61,62]
uPA/SCID mice reconstituted with PHH infected with HBV: [63,64] Viral assembly/encapsidation HAPs;
Bay 41-4109 (HAP compound)
Phenylpropenamides;
AT-61; AT-130 Inhibit HBV virion production by inappropriate assembly of capsids (no clinical data available yet)
Induces structural changes in HBV capsids; initiates virion assembly at wrong point in time (no clinical data available yet) Cell culture: [65,68–69]
HBV Cp(Cp149) expressed in Escherichia coli: [66,67]
Cell culture: [71]
HBV Cp(Cp149) expressed in E. coli: [73,74] HBsAg secretion/inhibition I: Nitazoxanide and its active metabolite, tizoxanide
II: Triazolo-pyrimidine inhibitors
III: NAP (Rep 9AC') I and II: selective inhibition of intracellular HBV replication and extracellular virus production; reduce levels of extracellular HBsAg, HBeAg, and levels of intracellular HBcAg in a dose-dependent manner. Synergistic activity in combination with 3TC or adefovir against HBsAg secretion in vitro
III: blocks HBsAg SVPs without affecting secretion of virions I: cell culture: [83]
II: cell culture: [78,80]
III: Proof of concept Phase I/II clinical trials on going. Interim results presented at the 48th annual meeting of EASL [182] Envelopment Glucosidase inhibitors Inhibition of glycosylation of envelope proteins in ER Study in animal model (WHV): [87,88]
Cell culture (Hep G2 cells): [86,89] cccDNA ZFNs
AAV vector platform to deliver ZFNs
DSS: CCC-0975 and CCC-0346
IFN-α and lymphotoxin-β receptor – APOBEC-dependent degradation of cccDNA
IFN-α Block transcription of cccDNA leading to inhibition of viral replication
Block conversion of rcDNA to cccDNA
IFN-α and lymphotoxin-β receptor activation upregulated APOBEC3A and APOBEC3B cytidine deaminases resp. resulting in cccDNA degradation
Epigenetic silencing of cccDNA; suppress HBV replication by transcriptional repression of cccDNA Cell culture (LMH cells + DHBV): [91]
Cell culture (Huh7/pTHBV2 HBV model system): [92,93]
Cell culture (DHBV infected PDH): [94]
Cell culture (HBV-infected cells, PHH, and dHepaRG cells): [95]
III: RNAi In vitro evidence of inhibition of viral replication I: cell culture: [97]
II: cell culture: [98,99]
III: cell culture and mice transfected with an HBV plasmid (small animal model): [102–104]
3TC: Lamivudine; cccDNA: Covalently closed circular DNA; CHB: Chronic HBV; Cp: HBV core promoter; DHBV infected PDH: Duck HBV-infected primary duck hepatocytes; dHepaRG: Differentiated HepaRG cells; DSS: Disubstituted sulfonamides; EAST: European Association for the Study of the Liver; HAP: Heteroaryldihydropyridine; HBcAg: Hepatitis B core antigen; HbeAg: Hepatitis B envelope antigen; HbsAg: Hepatitis B surface antigen; HBV: Hepatitis B virus; NTCP: Sodium taurocholate cotransporting polypeptide; PHH: Primary Human hepatocytes; ZFN: Zinc finger nuclease.
Table 2. Host factors Factors Targets Results Ref.
Innate immunity PRRs (TLRs) TLR-7 agonist: GS-9620 Suppression of HBV DNA in both serum and liver of HBV-infected chimpanzees; induction of IFN-α and cytokines
Phase I trials in CHB patients Animal study: [124]
Phase I study in healthy volunteers: [125]
Phase I trials in CHB (completed) [126,127] Cytokines IL-7
Recombinant human IL-7 (rhIL-7)-CYT 107
IL-21
Recombinant IL-21 Preclinical data:
Immunorestorative and vaccine adjuvant effect
Phase I–II clinical trial on CHB patients ongoing
Possible immunorestorative effect when used in combination with antivirals Small animal study (LCMV mice): [128,129]
Small animal study (LCMV mice): [134–135,137] Adaptive immunity Inhibitory T lymphocytes PD-1/PD-L1 Blockade of PD-1/PD-L1 interaction leading to restoration of T-cell function Small animal study (LCMV mice): [146]
GS-4774 (formerly GI-13020) Antigen-specific T-cell responses; tarmogens elicited HBV-specific T-cell responses ex vivo in samples collected from HVs and donors with CHB; induce both CD4+ and CD8+ T-cell responses in ex vivo model; induce HBV-specific T-cell response [172,183]
Phase I trial in HV completed: [184]
Phase II trial in CHB patients: [185] Adenovirus-based therapeutic vaccination TG1050 Stimulate polyfunctional, multispecific, robust and long-lasting T cells targeting multiple epitopes from three major viral proteins, expected to control the HBV replication and to elicit viral clearance [173] DNA vaccines Activate not only the T-cell responses specific to HBV but also natural killer cells Phase I clinical trial: [175]
Proof-of-concept study/CHB carriers: [176] T-cell peptide epitope vaccine String of 30 HBV-derived CTL epitopes linked to 16 Th epitopes presented to T cells by a large number of HLA molecules Mouse model: [177]
Novel Strategies to Achieve Sustained Virologic Remission新的战略,以实现持续病毒学缓解 Targeting the Virus针对病毒
Targeting Viral Entry. HBV is an enveloped virus with tropism to infect hepatocytes and viral entry is mediated through specific interactions of viral membrane proteins with cellular receptors. Targeting viral entry with receptor antagonists provides us with new opportunities to treat HBV. Recently, in vitro studies, using the primary hepatocytes from tupaia, Yan et al. reported a functional receptor for HBV, sodium taurocholate cotransporting polypeptide (NTCP).[60] NTCP is a sodium-dependent transporter for taurocholic acid, which is expressed at the basolateral membrane of hepatocytes and responsible for most Na+-dependent bile acid uptake in hepatocytes. In this regard, Myrcludex-B, a synthetic lipopeptide derived from pre-S1 domain of the HBV envelope protein, which specifically targets the NTCP has been shown to efficiently block HBV infection in in vitro [61,62] ]and in uPA/SCID mice reconstituted with human hepatocytes infected with HBV.[63,64] A Phase IIa clinical study in CHB patients, investigating the safety, tolerability and efficacy of multiple doses of Myrcludex B in comparison with the control group receiving standard therapy with NAs, is recently completed. Results are awaited. 针对病毒进入。 HBV是一种有包膜的病毒,嗜感染的肝细胞和病毒的进入是通过病毒膜蛋白与细胞受体的特异性相互作用介导的。针对病毒进入与受体拮抗剂为我们提供了新的机会来治疗乙肝。近日,在体外研究中,使用从树鼩,燕等人原代肝细胞。报道了功能性受体乙肝病毒,牛磺胆酸钠cotransporting多肽(NTCP)。[60] NTCP是钠依赖转运的牛磺胆酸,这是表示在肝细胞的基底膜,并负责大多数肝细胞的Na+依赖性胆汁酸的吸收。在这方面,Myrcludex-B,从HBV包膜蛋白,其特异性地靶向NTCP的前S1结构域衍生的合成脂肽已被证明能够有效地阻止HBV感染在体外[61,62],并在中uPA/ SCID小鼠重组,感染乙肝病毒的人肝细胞[63,64]一个IIa期临床研究慢性乙型肝炎患者,调查安全性,耐受性和多剂量Myrcludex乙的疗效与对照组接受标准疗法使用NAS相比,是最近完成。结果等待。
Targeting Viral Assembly/Encapsidation. HBV persistence and transmission require HBV replication, which depends on the assembly of a core particle composed of capsid protein (Cp), polymerase, and pregenomic RNA. Assembly is one of the critical steps in viral replication, which could be an attractive target for therapeutics. There are multiple classes of compounds discovered that could dysregulate or inhibit virion assembly and encapsidation. Heteroaryldihydropyrimidines are compounds that inhibit HBV virion production in vitro and in vivo by preventing encapsidation.[65,66] One of the most studied heteroaryldihydropyrimidine compounds is Bay 41-4109, which inhibits capsid formation, concomitant with a reduced half-life of the core protein. These drugs inhibit viral replication by inducing assembly inappropriately and, when in excess, by misdirecting assembly, decreasing the stability of normal capsids.[67–69] These compounds are also active against HBV mutants resistant to NAs.[70] 针对病毒装配/包壳。乙肝病毒的持久性和传输需要HBV的复制,这取决于衣壳蛋白(CP),聚合酶和RNA前基因组组成的核心颗粒的装配。组装是在病毒复制中的关键步骤,这可能是用于治疗的有吸引力的靶之一。有多个类化合物的发现可能dysregulate或抑制病毒组装和包壳的。 Heteroaryldihydropyrimidines是抑制HBV病毒颗粒的生产在体外和体内以防止壳体化化合物[65,66]其中研究最多的heteroaryldihydropyrimidine化合物是BAY41-4109,其抑制病毒衣壳的形成,伴随着芯的半衰期缩短蛋白质。这些药物抑制病毒复制由不适当感应组件,并且当过量时,通过误导装配,降低的正常衣壳的稳定性。[67-69],这些化合物也具有抗HBV变异株耐佐。[70]
Similarly, phenylpropenamides have also been shown to inhibit viral encapsidation, and are found to be active against 3TC-resistant strains.[70–72] Phenylpropenamides are shown to induce tertiary and quaternary structural changes in HBV capsids. AT-130 (phenylpropenamide derivative) has been shown to bind to a promiscuous pocket at the dimer–dimer interface that favors a unique quasiequivalent binding site in the capsid and can serve as an effective antiviral agent. It decreases viral production by initiating virion assembly at the wrong point in time, resulting in morphologically normal capsids that are empty and noninfectious.[73,74] Clinical efficacy of these compounds has not been reported yet and needs to be studied.
同样,phenylpropenamides也已显示出抑制病毒衣壳化,并且被发现是有效对抗3TC抗性株[70-72] Phenylpropenamides示以诱导乙型肝炎病毒的衣壳的三级和四级结构变化。 AT-130(phenylpropenamide衍生物)已经显示出结合于混杂口袋的二聚体二聚体界面有利于一个独特quasiequivalent结合位点中的衣壳,并可以作为一种有效的抗病毒剂。它减少病毒产生由引发病毒粒子装配在错误的时间点上,从而导致正常形态衣壳是空的和非感染性的。这些化合物[73,74]的临床疗效尚未见报道,需要进一步研究。
Targeting HBsAg Secretion. HBV persistence results from an ineffective antiviral immune response towards the virus. The exact mechanism by which HBV escapes immunity is poorly understood. The initial response to viral infection results in activation of innate immune responses such as the production of type I IFNs (IFN-α and IFN-β). 针对HBsAg的分泌。乙肝病毒的持久性的效果从对病毒无效的抗病毒免疫应答。由乙肝病毒逃逸免疫确切的机理知之甚少。病毒感染导致的激活先天免疫反应,如生产的I型干扰素(IFN-α和IFN-β)的初步反应。
Studies on HBV-infected chimpanzees demonstrated a complete lack of induction of type-1 IFN and IFN response genes during early stages of infection. It was recently shown that type-1 IFN responses are also lacking in acute HBV patients.[75,76] In this regard, the early stages of acute HBV are characterized by induction of IL-10 rather than type I IFN, accompanied by a temporary attenuation of natural killer (NK) cell and T-cell responses.[77] The suppression of innate immune response can also be mediated by direct interference of HBV antigens with host cells. High levels of HBsAg in the range of 400 μg/ml (0.4% of total serum protein) have been demonstrated in HBV infected patients[78–80] and are thought to play an important role in suppressing the HBV-specific immune response. In this regard, recent reports have suggested that HBsAg acts directly on dendritic cells to limit cytokine production.[81,82] Thus, control of HBsAg secretion could potentially enable its use with the therapeutic vaccine or as a combination therapy with NAs for the treatment of HBV. Several classes of drugs have been studied to reduce HBsAg secretion.[83] In vitro data showed that nonspecific antimicrobial nitazoxanide and its active metabolite, tizoxanide, reduced the levels of extracellular HBsAg, HBeAg, as well as the levels of intracellular HBcAg in a dose-dependent manner in vitro. Nitazoxanide was found to exhibit selective inhibition of intracellular HBV replication and extracellular virus production in cell cultures, and synergistic activity in combination with lamivudine or adefovir against HBV.[83] Recently, a series of novel triazolo-pyrimidine inhibitors of HBsAg secretion was identified using the HBV-expressing cell line HepG2.2.15, through high-throughput screening. The parent compound was shown to not be an inhibitor of viral genomic replication but rather a specific inhibitor of HBV envelope secretion. These triazolo-pyrimidine derivatives were also active in inhibiting HBsAg secretion of HBV variants that are resistant to current NAs.[78,80] The exact mechanism of action for these compounds is still under investigation, and it is unclear at the moment if the reduction of HBsAg secretion would be able to enhance HBV specific immunity in vivo.
在感染的早期阶段研究HBV感染的黑猩猩表现出完全缺乏诱导1型干扰素和干扰素反应基因的。据最近表明,1型干扰素的反应也缺乏在急性乙型肝炎患者[75,76]在这方面,急性HBV的早期阶段的特征是感应IL-10,而不是I型IFN的,伴随着自然杀伤(NK)细胞和T细胞应答的临时衰减。[77]的先天免疫反应的抑制,也可以通过HBV抗原与宿主细胞直接干扰介导的。在400微克/毫升(总血清蛋白的0.4%)的范围内的高水平的HBsAg已经证实在HBV感染的患者[78-80],并且被认为在抑制HBV特异性免疫应答中发挥重要作用。在这方面,最近的报告表明,乙肝表面抗原直接作用于树突状细胞,以限制产生细胞因子[81,82]因此,乙肝表面抗原分泌的控制可能使其与治疗性疫苗或作为组合疗法使用NAS用于治疗用途乙肝病毒。几类药物进行了研究,以减少HBsAg的分泌[83]体外数据表明,非特异性抗菌硝唑尼特和其活性代谢物,tizoxanide,降低细胞外的HBsAg和HBeAg的水平,以及细胞内的HBcAg的水平以剂量依赖的方式在体外。硝唑尼特,发现表现出细胞内HBV复制和细胞外病毒生产细胞培养物,并在用拉米夫定或阿德福韦抗HBV组合的协同活性的选择性抑制作用。[83]最近,一系列的HBsAg分泌的新型三唑并嘧啶抑制剂,使用鉴定HBV的表达细胞系HepG2.2.15,通过高通量筛选。母体化合物被证明是没有病毒基因组复制的抑制剂,而是HBV包膜分泌的特异性抑制剂。这些三唑并嘧啶衍生物也积极地抑制乙肝病毒的变种能够抵抗当前NAS HBsAg的分泌[78,80]的行动对这些化合物的确切机制仍在调查中,目前还不清楚目前是否减少HBsAg的分泌就能够提高HBV特异性免疫的体内。
The HBV genome is an extremely compact structure (~3 kb), which encompasses four overlapping ORFs encoding for viral polymerase/reverse transcriptase; the capsid-forming core protein; three envelope proteins called large (LHBs), middle (MHBs) and small hepatitis B surface antigens; and the regulatory X protein.
HBV基因组是一种结构非常紧凑的(〜3 kb的),它包括四个重叠的ORF编码病毒聚合酶/逆转录酶;衣壳形成核心蛋白; 3包膜蛋白称为大(LHBs的),中间(MHBs)和小乙型肝炎表面抗原;和监管X蛋白。
The HBV virion is a double-shelled sphere with an inner nucleocapsid and an outer lipoprotein envelope. In addition to infectious virion, HBV produces two other types of particles (subviral empty envelope particles and subviral naked capsid particles). The three envelope proteins are present in different proportions in three types of HBV particles. Noninfectious subviral particles (SVPs) share antigenic features of the virus envelope and are thought to presumably act as a decoy for the immune system.[84] In the past, it was believed that budding of infectious virions and SVPs used the same pathway. However, recent reports suggest that budding of infectious virions depends on functions of the multivesicular body pathway.[84]
乙肝病毒体是双壳球体具有内部核衣壳,外脂蛋白包膜。除了感染病毒,乙肝病毒产生两种其他类型的颗粒(亚病毒空信封颗粒和亚病毒衣壳裸体颗粒)。三种包膜蛋白是存在于三种类型的HBV颗粒的比例不同。非感染性亚病毒颗粒(的SVP)共享病毒包膜的抗原性特征,并且被认为大概是作为诱饵对免疫系统[84]在过去,人们认为感染性病毒粒子和SVP的出芽所使用的相同的途径。然而,最近的报告表明,感染性病毒颗粒的出芽取决于多泡体途径的功能。[84]
Nucleic acid polymer, amphipathic oligonucleotide is currently undergoing a proof-of-concept trial in patients with CHB. Rep 9AC' blocks the secretion of HBsAg SVPs without affecting the secretion of infectious virion (which is line with recent studies, suggesting that SVPs and infectious virion use different pathway for budding). Patients treated with REP 9AC' (nucleic acid polymer) cleared HBsAg from serum and some achieved seroconversion. Updated interim results were presented at the 49th annual meeting of the European Association for the Study of the Liver held in Amsterdam (The Netherlands). Patients who had cleared HBsAg from their blood with REP 9AC' monotherapy were subjected to combination treatment with REP 9AC' and either PEG-IFN or Thymosin-α (compound with immunomodulatory activity). Further increases in anti-HBV antibodies were observed in all patients with combination treatment. Further updates on this trial are awaited.
核酸聚合物,两亲性寡核苷酸目前正在进行概念验证的试验CHB患者。REP9AC'块的HBsAg SVP的,而不会影响感染病毒的分泌物(这是符合最近的研究,提示的SVP和感染病毒使用不同的途径萌芽)的分泌。与REP9AC'(核酸聚合物)治疗的患者中清除的HBsAg从血清和一些实现血清转化。最新中期结果发表在欧洲协会的肝脏在阿姆斯特丹(荷兰)举行的研究第49届年会。谁曾清除乙肝表面抗原从血REP9AC患者的单药治疗经受结合治疗REP9AC',要么聚乙二醇干扰素和胸腺肽α(与免疫调节活性化合物)。在抗乙肝病毒抗体进一步增加,观察所有患者的综合治疗。在本次试验的进一步更新等待。
Targeting Envelopment. All three HBV envelope proteins share the same S domain and contain N-linked glycosylation at amino acid 146 within the S domain. MHBs and LHBs contain the pre-S2 domain, while N-terminal of LHBs protein contains the pre-S1 domain. Envelopment of hepatitis B core particle (capsids) containing the HBV genome depends on the interaction of nonglycosylated pre-S sequences facing to the cytosol with defined regions on the core particle. The transport of the enveloped virus particles and SVPs particles may indeed depend on glycosylation and processing of the viral MHBs glycoprotein. 针对包络。所有三个HBV包膜蛋白共享相同的S域和包含在S域中的N连接的糖基化的氨基酸146。 MHBs和LHBs的含有前S2结构域,而LHBs的蛋白质的N末端含有前S1结构域。含有HBV基因B型肝炎核心颗粒(衣壳)的包络取决于朝向胞质溶胶与在核粒子定义的区域非糖基化前S序列的相互作用。的包膜病毒粒子和的SVP粒子的运输可能确实依赖于糖基化的病毒MHBs糖蛋白的加工。
Glucosidase inhibitors inhibit viral morphogenesis and infectivity, most likely by inhibition of glycosylation of envelope protein in ER.[85–89] This approach is important to disrupt the development of HBV envelope, thereby generating quasispecies, that is defective in its ability to bind target cells and establish infection. This is another promising approach to target HBV.
葡萄糖苷酶抑制剂抑制病毒的形态发生和感染性,最有可能是通过抑制雌激素受体的包膜蛋白的糖基化[85-89]这种方法是很重要的,破坏HBV包膜的发展,从而生成准种,即有缺陷的在其结合靶的能力细胞并建立感染。这又是一个很有前途的方法针对乙肝病毒。
Targeting cccDNA. Current NA-based treatment can block the replication and formation of new cccDNA, but existing cccDNA in already infected cells is not affected directly by current therapies and it has a long half-life (33–50 days). The viral cccDNA in the nucleus serve as a store of viral escape variants generated by the error-prone viral polymerase and escape mutants conferring drug-resistance, which can cause drug resistance or viral rebound upon cessation of treatments. 针对cccDNA的。当前NA为基础的治疗可阻断在已感染的细胞中复制并形成新的cccDNA,但现有的cccDNA不是由当前疗法的直接影响,它具有长的半衰期(33-50天)。病毒的cccDNA在细胞核中作为由易错的病毒聚合酶产生的病毒逃避变体的一个商店和逃避突变体赋予耐药性,一旦停止治疗,可导致耐药性或病毒反弹。
Inactivation/Elimination/Degradation of cccDNA. A recently developed therapeutic approach that directly targets HBV cccDNA within cells is by use of zinc-finger nucleases (ZFNs). Zinc finger proteins can be used to block the transcription of cccDNA. ZFNs have proved as one of most versatile and effective classes of gene targeting reagents in recent years. ZFNs have separate DNA-binding and DNA-cleavage domains. For more detail on ZFN engineering please read reference.[90] cccDNA的灭活/消除/降解。最近开发的治疗方法是直接针对乙肝病毒cccDNA的细胞中是通过使用锌指核酸酶(ZFN)的。锌指蛋白可以用于阻断cccDNA的转录。的ZFN已被证明是最通用的,有效的类基因靶向试剂近年来之一。的ZFN有独立的DNA结合和DNA切割结构域。有关ZFN技术详细请阅读参考。[90]
There is evidence in vitro that ZFPs can be used to specifically target the cccDNA of duck HBV infection and inhibit viral transcription and replication.[91] Expression of the ZFPs in LMH cells undergoing the DHBV viral lifecycle resulted in decreased expression of viral RNA and protein expression compared with the empty vector control, without any apparent toxicity effects. In addition, the production of viral particles was also decreased in the presence of the expressed ZFPs.[91]
有体外的锌指蛋白可用于特异性靶向的鸭HBV感染的cccDNA和抑制病毒转录和复制的证据。的锌指蛋白的[91]在LMH细胞经历DHBV病毒生命周期中的表达导致降低的病毒RNA和蛋白表达与空载体对照的表达相比,没有任何明显的毒性作用。此外,所生产的病毒颗粒也被降低,在表达锌指蛋白的存在[91]。
ZFNs are able to cleave HBV DNA in hepatoma cells in vitro. However, it raises clinical challenge of off target effects and specifically delivering ZFNs to the liver of infected subjects.[92] This challenge could be overcome by the application of vector platform for delivery. Studies using adeno-associated virus as a vector platform is being explored to deliver designer nucleases to target cells. Adeno-associated virus vectors were found to be safe in clinical and preclinical applications.[93]
的ZFN能够切割HBV DNA在肝细胞瘤细胞在体外。然而,它提出的脱靶效应和特异性递送的ZFN对受感染的受试者的肝脏临床挑战。[92]这种挑战可以通过向量平台的应用递送克服。用腺相关病毒作为载体平台的研究正在探讨以提供设计者核酸至靶细胞。腺相关病毒载体被认为是安全的,临床和临床前应用中。[93]
Recently Cai et al. identified two structurally related disubstituted sulfonamides (DSS), termed CCC-0975 and CCC-0346, which were confirmed as inhibitors of cccDNA production, with low micromolar EC50 in cell culture. The author demonstrated that DSS was able to synchronously reduce levels of HBV cccDNA and its putative precursor, deproteinized relaxed circular DNA (DP-rcDNA), without directly inhibiting HBV DNA replication in cell culture or reduction in viral polymerase activity. However, DSS compounds did not promote the intracellular decay of HBV DP-rc DNA and cccDNA, suggesting that the compounds interfere primarily with rcDNA conversion into cccDNA. In addition, CCC-0975 was shown to reduce cccDNA biosynthesis in duck HBV-infected primary duck hepatocytes.[94]
最近Cai等。确定了两个结构相关取代磺胺类(DSS),被称为CCC-0975和CCC-0346,它被确认为cccDNA的产生的抑制剂,在细胞培养小微摩尔EC50。作者表明,决策支持系统能够同步降低乙肝病毒cccDNA的和公认的前体的水平,去蛋白松弛环状DNA(DP-rcDNA抑制),不直接抑制乙肝病毒DNA复制的细胞培养或减少病毒聚合酶的活性。然而,决策支持系统的化合物没有促进HBV DP-RC DNA和cccDNA的细胞内的衰变,这表明该化合物干扰主要与rcDNA抑制转化的cccDNA。此外,CCC-0975被证实可以降低cccDNA的生物合成鸭乙型肝炎病毒感染的原发性肝细胞的鸭子。[94]
Another very recent study demonstrated APOBEC-dependent degradation of HBV cccDNA induced by IFN-α and lymphotoxin-β receptor on HBV-infected, differentiated HepaRG (dHepaRG) cells and primary human hepatocytes. HBV core protein mediated the interaction with nuclear cccDNA, resulting in cytidine deamination, apurinic/apyrimidinic site formation, and finally cccDNA degradation that prevented HBV reactivation.[95]
另一个非常最近的研究表明HBV cccDNA的诱导IFN-α和淋巴毒素-β受体在HBV感染,分化的HepaRG(dHepaRG)细胞和人原代肝APOBEC依赖性降解。乙肝病毒核心蛋白介导的核cccDNA的相互作用,导致胞苷脱氨,脱嘌呤/脱嘧啶位点的形成,并最终降解的cccDNA,阻止乙肝病毒再激活。[95]
Epigenetic Silencing of cccDNA. Another approach of targeting cccDNA is by epigenetically silencing or transcriptional repression. There is evidence of epigenetic silencing of cccDNA by IFN-α in cell culture and in humanized mice. IFN-α was shown to suppress HBV replication by targeting the epigenetic control of cccDNA function and transcription.[96] cccDNA的表观遗传沉默。靶向的cccDNA的另一种方法是通过表观遗传学上沉默或转录抑制。有cccDNA的在细胞培养物和人源化的小鼠后生沉默由IFN-α的证据。 IFN-α所表现出cccDNA的目标函数和转录的表观遗传调控来抑制HBV的复制。[96
Targeting Viral mRNA. Viral mRNA can be directly targeted using antisense oligonucleotides, ribozymes, or RNAi. In vitro data exist that show that HBV transcript levels can be reduced by using antisense oligonucleotides,[97] hairpin ribozymes[98] or hammerhead ribozymes using lentiviral vector for delivery.[99] 针对病毒mRNA。病毒mRNA可以用反义寡核苷酸,核酶或RNAi直接针对性。体外数据使用慢病毒载体交付存在表明,乙肝病毒转录水平可以用反义寡核苷酸降低,[97]发夹核酶[98]或锤头状核酶。[99]
RNAi is one of the fastest moving fields in modern biotechnology. RNAi is a process by which small interfering RNA molecules induces gene silence at post-transcriptional level to effectively knock down the expression of genes of interest. In mammalian cells, it can be used specifically to target the degradation of cellular mRNA.[100] Due to extensive use of ORFs with the DNA genome in HBV, multiple HBV RNAs will make the virus susceptible for RNAi.[101] Several studies have shown that viral mRNA and HBV replication can be inhibited by using RNAi in cell cultures and in mice models.[102–105] McCaffrey et al. showed that RNAi could be applied to inhibit production of HBV replicative intermediates in cell culture and in immunocompetent and immunodeficient mice transfected with a HBV plasmid. RNAi was able to inhibit all the steps of HBV replication that occur in cell culture and in mice. They found four separate lines of evidence to establish that RNAi substantially inhibited HBV in mice: RNAi expression significantly reduced secreted HBsAg in serum; HBV RNAs were substantially reduced in mouse liver; HBV genomic DNA was reduced to undetectable levels in mouse liver; and the number of cells staining for HBV core antigen (HBcAg) was substantially decreased.[103] RNA-i could be used as a novel therapeutic approach, however, several challenges such as efficient delivery in vivo, RNA instability and off target effects exists and needs to be overcome before it can be exploited in treatment of CHB patients.
RNAi是在现代生物技术的移动最快的领域之一。 RNAi是一个过程,小干扰RNA分子诱导基因沉默在转录后水平上,有效地击倒感兴趣的基因的表达。在哺乳动物细胞中,可以具体地用于靶向细胞mRNA的降解。[100]由于采用了大量与HBV DNA的基因组中的ORF,复合HBV的RNA将会使病毒易感于RNAi的[101]许多研究都表明,病毒mRNA和HBV复制可以通过在细胞培养物和小鼠模型使用的RNAi来抑制。[102-105] McCaffrey的等表明,RNAi技术可应用于抑制生产的HBV复制中间体在细胞培养和转染了HBV质粒免疫和免疫缺陷小鼠。 RNA干扰是能够抑制乙型肝炎病毒复制的所有发生在细胞培养物和小鼠中的步骤。他们发现的证据,四个单独的行建立了RNA干扰对小鼠显着抑制乙肝病毒:RNAi表达显著降低血清中HBsAg的分泌;乙肝病毒的RNA被大幅降低小鼠肝脏; HBV基因组DNA减少到检测不到的水平在小鼠肝脏;和细胞染色的HBV核心抗原(HBcAg的)的数量基本上减少。[103]的RNA-i的可作为一种新的治疗方法,然而,一些挑战,如高效的递送体内,RNA不稳定和脱靶效应的存在并且需要被克服之前,它可以在治疗慢性乙型肝炎患者的被利用。 作者: StephenW 时间: 2014-9-8 16:46
Targeting the Host针对主机
Recovery from acute HBV infection is associated with robust innate and adaptive immune responses. Innate immune response is the first line of defense against viral infections and results in production of type I IFN, which leads to suppression of viral replication, mediation of NK cell-mediated killing of viral infected cells, and supports the efficient maturation and site recruitment of adaptive immunity through production of proinflammatory cytokines and chemokines.[59,106] These IFN enhance the first defense against viral infections and modulate both innate and adaptive immune cells.[77,107] The principal producers of type I IFN are the plasmacytoid dendritic cells (pDC) that respond to viruses and other pathogens primarily through the recognition of pathogen-associated molecular patterns by two intracellular TLRs: TLR7 and TLR9.[77,108] Triggering of TLR leads to activation of pDC and production of high levels of type I IFNs, along with the release of other cytokines, including TNF-α, IL-6 and cell surface costimulatory molecules. pDC also activate NK cells and T lymphocytes, allowing further priming and regulation of antiviral immunity.[107,109–110] Efficient priming of the adaptive immune system causes functional maturation and expansion of distinct B- and T-cell clones, which are able to specifically recognize the infectious agents. This process leads to control of infection and generates a memory response that increases the host ability to block subsequent infections with the same pathogens.[59]
恢复从急性HBV感染与鲁棒先天和适应性免疫反应相关联。先天免疫应答是抵御病毒感染,并导致生产的I型IFN,这导致抑制病毒复制,NK细胞介导的杀伤病毒感染细胞的调解的第一行,并且支持高效的成熟和现场招聘通过促炎性细胞因子和趋化因子的适应性免疫。[59106]这些干扰素增强抗病毒感染的第一道防线,并调节这两个先天和适应性免疫细胞。[77107]式的主要生产者I型IFN是浆细胞样树突细胞(PDC),该主要是通过由两个胞内的TLR识别病原体相关的分子模式响应病毒和其他病原体:TLR7和TLR9[77108]触发的TLR导致活化的pDC和生产高水平予IFN的类型,随着释放的其他细胞因子,包括TNF-α,IL-6和细胞表面的共刺激分子。的pDC也激活NK细胞和T淋巴细胞,从而进一步引发和抗病毒免疫的调节[107,109-110]适应性免疫系统的高效引发导致功能性成熟和扩张的鲜明的B和T细胞克隆,其能够特异性识别该传染性病原体。这个过程导致感染的控制,并产生记忆反应,增加阻止继发性感染具有相同的病原体的宿主的能力。[59]
In CHB patients, HBV is associated with blunting of innate and adaptive immune responses. Hence, strategies that would augment innate immune responses may also enhance adaptive anti-HBV immunity. Several studies have demonstrated that expressions of TLRs (TLR 2, TLR 3, TLR4, TLR7 and TLR9) were decreased in CHB patients.[111–114] In this regard, HBV interferes with TLR2, 7 and 9 signaling, which are considered to play an important role in the control of infection and elimination of virally infected cells.
慢性乙型肝炎患者,乙肝病毒与钝化的先天免疫和适应性免疫反应有关。因此,战略,将增强先天免疫反应也可增强自适应抗乙肝病毒的免疫力。几项研究已经表明,Toll样受体的表达(TLR2,TLR3,TLR4,TLR7和TLR9)分别降低慢性乙型肝炎患者[111-114]在这方面,HBV干扰TLR2,7,9信令,这被认为是玩在感染的病毒感染的细胞的控制和消灭具有重要作用。
TLR Agonists. Studies have shown that when HBV transgenic mice were injected with ligands specific for TLR 2–9, liver of these HBV transgenic mice produce IFN-α, -β and -γ to inhibit HBV replication, suggesting that HBV replication can be controlled by the activation of innate immune response in the liver.[115] The inhibition of HBV replication was accomplished at a post-transcriptional level by suppressing the assembly or stability of HBV RNA-containing capsids.[115,116] These provide evidence that TLR activation directly inhibits HBV replication.[117,118] However, HBV somehow evades innate recognition by TLRs as a strategy to escape innate immune response by its ability to disrupt TLR expression and inhibit TLR signaling cascades.[117,118] It has been reported that the expression of TLRs in hepatocytes and other cells is downregulated in the presence of various HBV viral products.[112,119–123] Although HBV circumvents endogenous type I IFN pathways, it is plausible that the use of exogenous IFN induction using the TLR7 agonist may reinstate the IFN-α response. When combined with a strategy that results in maximal suppression of HBV replication in vivo using NAs, exogenous IFN stimulation via TLR agonists may result in development of protective immunity. Several studies have shown that long-term suppression of HBV using NA results in partial reconstitution of adaptive immunity. In this regard, an adjuvant therapy using TLR agonist may able to accelerate this process of immune reconstitution and HBV clearance. First, TLR agonists are available as oral compounds enabling rapid uptake by the liver. Second, they may allow being combined along with other NAs as a single pill. Finally, similar to injected IFN, TLR agonists induce IFN production, triggering the production of cytokines to facilitate intracellular communication and cellular trafficking. However, through the use of TLR agonists this antiviral state can be induced at the liver, eliminating the adverse events associated with systemic innate immune activation. Toll样受体激动剂。研究表明,当HBV转基因小鼠注射特异性针对TLR2-9配位体,这些HBV转基因小鼠的肝脏中产生的IFN-α,-β和-γ抑制乙型肝炎病毒复制,这表明HBV复制可以通过激活控制在肝脏中的先天免疫应答。[115] HBV复制的抑制是通过抑制HBV RNA的含衣壳的组装或稳定性来实现在转录后水平。[115116]这些提供的证据表明,TLR活化直接抑制HBV复制[117118]然而,HBV某种方式通过TLR的逃避先天识别作为一个策略,以逃避通过其扰乱TLR表达和抑制TLR信号传导级联的能力先天免疫应答[117118]已经报道了Toll样受体在肝细胞和其他的表达细胞被下调中的各种B型肝炎病毒的产品的存在。[112,119-123]尽管HBV绕过内源I型IFN途径,它是合理的,使用用TLR7激动剂外源性干扰素的诱导可能恢复的IFN-α应答。当与一种策略使用NAS其导致HBV复制的最大值抑制在体内结合,经由TLR激动剂外源性IFN刺激可导致保护性免疫的发展。几项研究已经表明,HBV的使用NA长期抑制导致获得性免疫的部分重构。在这方面,使用的TLR激动剂佐剂疗法可能能够加速免疫重建和HBV间隙的这一过程。首先,Toll样受体激动剂可作为口服化合物由肝脏能够快速吸收。其次,他们可能会被允许与其他NAS,作为一个单一的药丸结合一起。最后,类似于注射干扰素,TLR激动剂诱导干扰素的产生,引发细胞因子的产生,以促进胞内通信和细胞运输。然而,通过使用的TLR激动剂此抗病毒状态,可在肝脏引起的,消除了与全身先天性免疫的激活相关的不良事件。
Lanford et al., investigated the effects of immune activation with GS-9620, an orally administered agonist of TLR-7, in chimpanzees chronically infected with HBV. GS-9620 was administered to chimpanzees every other day for 4 weeks at 1 mg/kg and, after a 1-week rest, for a second cycle of 4 weeks at 2 mg/kg. TLR-7 agonists induced prolonged suppression of HBV DNA in both the serum and liver.[124] GS-9620 administration induced the production of IFN-α and various cytokines and chemokines. In addition, it activated all lymphocyte subsets to induce interferon stimulated genes (ISGs).[124] HBV DNA was reduced, in addition to serum levels of HBsAg, HBeAg and HBV antigen positive hepatocytes. However, an increase in liver enzymes and immune cell infiltration was observed during the period of decrease in both intrahepatic and serum viral load.
兰福德等人,研究了免疫激活作用和GS-9620,TLR-7的口服给药的激动剂,在慢性感染HBV的黑猩猩。 GS-9620在1毫克/公斤为2毫克/千克给予黑猩猩隔日4周,后1周的休息,为4周的第二轮。 TLR-7激动剂诱导的HBV DNA在血清和肝脏两者的延长抑制[124] GS-9620施用诱导产生IFN-α和各种细胞因子和趋化因子。此外,激活的所有淋巴细胞亚群诱导干扰素刺激基因(的ISG)[124]的HBV DNA的减少,除了对HBsAg,HBeAg和HBV抗原阳性肝细胞中的血清水平。然而,在减少在肝内和血清病毒载量的期间观察到的增加的肝酶和免疫细胞浸润。
In early studies to investigate the safety, tolerability, pharmacokinetics and pharmacodynamics of oral GS-9620 in healthy volunteers, oral doses (single dose of 0.3, 1, 2, 3, 4, 6, 8 or 12 mg) were well absorbed and well tolerated. Adverse events associated with IFN treatment were seen in subjects who received the 8 or 12 mg dose and serum IFN-α was only detected at these doses although activation or ISGs were seen at doses ≥2 mg.[125] Two Phase I clinical trials of GS-9620 on CHB patients are completed. One in virologically suppressed subjects with CHB, and other treatment naive subjects with CHB.[126,127] It will be interesting to know the results of these trials.
在早期的研究中,调查了安全性,耐受性,药代动力学和口服GS-9620在健康志愿者中的药效学,口服剂量(0.3,1,2,3,4,6,8或12毫克单一剂量)被很好地吸收和良好容忍。与干扰素治疗相关的不良事件主要出现在谁收到的8或12毫克的剂量,血清IFN-α仅在这些剂量检测,虽然激活的ISG或者被发现在剂量≥2毫克。[125]两个I期临床试验受试者GS-9620对慢性乙肝患者完成。一位在病毒学抑制受试者慢性乙型肝炎和慢性乙肝等治疗天真的科目。[126127]这将是有趣的知道这些试验的结果。
Cytokines. The use of cytokines as immunomodulatory therapy for the treatment of chronic viral infections has been extensively studied. Most promising candidates that may be beneficial in CHB patients are IL-7 and IL-21.[128–130] 细胞因子。使用细胞因子作为免疫调节治疗慢性病毒感染的治疗中已被广泛研究。最有希望的候选人,可能在慢性乙型肝炎患者是有益的,IL-7和IL-21。[128-130]
IL-7. IL-7 is absolutely essential for primary T-cell development and probably has an important role in the normal B-cell development process. IL-7 also plays a role in the development of some dendritic cell (DC) subsets. IL-7-mediated signaling in DCs has been shown to regulate peripheral CD4+ T-cell homeostasis.[131] IL-7 therapy for CHB patients would be to enhance, rejuvenate and restore immune response to HBV. Preclinical data generated from numerous model systems have shown that IL-7 has potent immunorestorative effects, as well as vaccine adjuvant effects and beneficial effects in the setting of adoptive cell therapy. CYT107 is a second-generation recombinant human IL-7 product made by Cytheris SA via a recombinant mammalian cell culture system.[132] A Phase I/II randomized, open labeled, controlled, dose-escalation study of repeated administration of recombinant human IL-7 (CYT107), in combination with standard antiviral treatment and vaccination in HBeAg-negative CHB patients is ongoing.[133]
IL-7。 IL-7是用于初级T-细胞的发展是绝对必要的,可能具有在正常B细胞发育过程中起重要作用。 IL-7也起着一些树突状细胞(DC)的子集的发展中的作用。 DC中的IL-7介导的信号已被证明是调节外周血CD4 + T细胞稳态。[131]的IL-7治疗慢性乙型肝炎患者会提升,恢复活力和恢复对HBV的免疫应答。从众多的模型系统中产生的临床前数据表明,IL-7具有强效immunorestorative效果,以及疫苗的佐剂效应,在过继性细胞治疗的设定的有益作用。 CYT107是第二代重组人IL-7产品通过重组哺乳动物细胞培养系统,通过Cytheris SA进行。[132]一个I / II期随机,开放标记,对照,剂量递增的重组人白细胞介素重复给药的研究-7(CYT107),符合标准的抗病毒治疗和疫苗接种HBeAg阴性CHB患者结合正在进行中。[133]
IL-21. IL-21 mediates an important function in the induction and maintenance of effector CD8+ T cells. Several studies done on chronic lymphocytic choriomeningitis virus (LCMV) infection in mice have shown that IL-21 signaling is required to maintain a functional pool of effector CD8+ T cells. Mice deficient in IL-21 or IL-21R show a progressive decline in the number of virus-specific polyfunctional effector CD8+ T cells, which correlates with poor viral control.[134–136] In CHB infection, IL-21 was shown to be critical in promoting immune responses that can control infection in mice.[137]
IL-21。 IL-21介导的诱导和维持效应CD8 + T细胞的重要功能。在小鼠上进行的慢性淋巴细胞性脉络丛脑膜炎病毒(LCMV)感染的几项研究表明,IL-21信号传导是必需的,以保持效应CD8 + T细胞的功能池。缺陷的小鼠中,IL-21或IL-21R示出了病毒特异的多官能效应CD8 + T细胞的数量,这与病毒对照差逐渐下降。[134-136]在慢性乙型肝炎感染,IL-21被证实为促进免疫反应,可以在小鼠中控制感染的关键。[137]
In a longitudinal study of CHB patients, it was demonstrated that patients treated with antivirals with complete suppression had significantly higher levels of serum IL-21 than those who did not.[138] High serum IL-21 concentrations were also predictive of HBeAg seroconversion, a clinically important outcome associated with control of HBV infection.[138] Recombinant IL-21 is a new immune modulator currently undergoing Phase I and II testing in cancer patients.[139] Use of recombinant IL-21 for CHB might be a promising approach as a combination therapy for CHB patients receiving NAs.
慢性乙型肝炎患者的纵向研究中,我们证实了抗病毒药物完全抑制治疗的患者血清中IL-21的显著更高水平比那些谁没有。[138]高血清中IL-21浓度也预测HBeAg血清学转换的,与HBV感染控制相关的重要的临床结果。[138]重组IL-21是一种新型的免疫调节剂目前正在进行一期和二期试验的癌症患者。[139]用重组IL-21对慢性乙型肝炎可能是一种很有前途的方法作为联合治疗用于接收新来港慢性乙型肝炎患者。
Programmed Death-1/Programmed Death Ligand-1. Chronic and persistent HBV infection is associated with weak and functionally impaired immune response. The persistent exposure to viral antigens leads to virus-specific CD4 and CD8 cell dysfunction or deletion, and with prolonged exposure leads to exhaustion of T-cell response. 程序性死亡1/程序性死亡配体1。慢性HBV持续感染与弱,功能受损的免疫反应有关。持续暴露于病毒抗原导致病毒特异性的CD4和CD8细胞的功能障碍或缺失,并且与长期暴露导致的T细胞应答的疲劳。
Recent data suggest these virus specific T cells hyperexpress the PD-1 molecule and interaction between programmed death-1 (PD-1) receptor on lymphocytes and its ligand programmed death ligand-1 (PD-L1)/2 plays an important role in T-cell exhaustion.[140–145] In vitro and in vivo data suggest that inhibition of PD-1/PD-L1/2 ligand, which blocks the engagement of PD-1 with its ligand (PD-L1/2), has shown improvement in the antiviral functions of these T cells.[146–152] Studies have shown that HBV-specific T cells express PD-1 at a higher level than other T cells and exhibit exhaustive functionality as determined by cytokine secretion.[140–141,143–144] Attempts have been made to reverse the PD-1–PD-L interaction in vitro to rejuvenate HBV-specific immunity. In this regard, in vitro blockade of PD-1/PDL1 in a woodchuck hepatitis model with chronically infected WHV showed restoration of T-cell function.[153] Moreover, in vivo blockade of PD1/PD-L1 along with therapeutic vaccination and antiviral NA treatment in persistently WHV-infected woodchucks showed that this combination resulted in potent antiviral effect and improved function of woodchuck hepatitis core antigen-specific CD8 T cells.[154]
最近的数据表明这些病毒特异性T细胞hyperexpress对PD-1分子和程序性死亡-1(PD-1)受体之间相互作用对淋巴细胞和其配体程序性死亡配体-1(PD-L1)/ 2起着T中起重要作用β细胞衰竭。[140-145]在体外和体内数据表明,抑制PD-1/ PD-L1/ 2的配体,其阻断PD-1与其配体(PD-L1/ 2)的卡合,具有在这些T细胞的抗病毒功能的明显的改善。[146-152]研究表明,HBV特异性T细胞表达的PD-1在较高的水平比其它的T细胞,并表现出如通过细胞因子的分泌来确定详尽的功能[140- 141,143-144]已经尝试扭转体外对PD-1-PD-L相互作用,以恢复HBV特异性免疫。在这方面,在土拨鼠肝炎模型与慢性感染WHV体外阻断PD-1 / PDL1的显示恢复的T细胞的功能。[153]此外,在体内阻断PD1 / PD-L1的同时治疗性疫苗和抗病毒NA治疗坚持不懈WHV感染的土拨鼠表明,这种组合导致了有效的抗病毒效果,并改善土拨鼠肝炎核心抗原特异性CD8 T细胞的功能。[154]
Fisicaro et al. examined the role of T-cell exhaustion in the pathogenesis of chronic HBV infection in patients with CHB. They compared phenotype and function of intrahepatic and circulating HBV-specific T cells, and effect of PD-1/PD-L1 blockade. Results showed that intrahepatic HBV-specific CD8 cells express higher PD-1 and lower IL-7 receptor, CD127 levels. PD-1/PD-L1 blockade led to T-cell restoration, both in the periphery and in the liver, with better functional improvement among intrahepatic T cells.[155]
Fisicaro等。研究了T细胞耗竭的作用在慢性HBV感染患者的慢性乙肝的发病机制。他们比较了表型和肝内和循环HBV特异性T细胞的功能,和PD-1 / PD-L1阻断的效果。结果表明,肝内HBV特异性CD8细胞表达更高的PD-1和下部的IL-7受体,CD127的水平。 PD-1/ PD-L1阻断导致T细胞的修复,无论是在周边和在肝脏,肝内T细胞之间更好的功能改善。[155]
Intravenous PD-1 and PD-L1 antibodies have been tested on patients with advanced cancer. Antibody-mediated blockade of PD-L1 induced durable tumor regression and prolonged stabilization of disease in patients with advanced cancers.[156] However, use of anti-PD-1 antibody was associated with a relatively high frequency of grade 3 or 4 adverse events (adverse events occurred in 14% of patients including three deaths from pulmonary toxicity).[157] Utilization of PD-1 blockade as a therapeutic modality for CHB patients is being investigated. This approach is unique and offers an excellent opportunity to revive exhausted T cells in CHB, thereby allowing restoration of adaptive immunity against HBV and offers a fair chance of achieving sustained virologic remission. Utilizing any approach to block central immunoregulatory mechanisms is associated with unsuspected complications. These constitute a risk profile that may not be acceptable to otherwise healthy CHB patients as compared with terminally ill cancer patients. Use of anti-PD-1 antibodies offers the best hope of blocking PD-1/L12 interaction, as PD-L1 block would still allow PD-1–PDL2 interaction, however, anti-PD-1 antibodies have been associated with most adverse events and may require further studies prior to application as a therapeutic agent for CHB.
静脉注射,PD-1和PD-L1的抗体已经过测试,在晚期癌症患者。 PD-L1的抗体介导的封锁引起的耐用肿瘤消退和疾病治疗晚期癌症的长期稳定。[156]然而,使用抗PD-1抗体为3级比较高的频率或4级不良事件有关(发生在患者包括肺毒性三人死亡14%的不良事件)的PD-1阻断的治疗方法对慢性乙型肝炎患者[157]利用正在调查中。这种方法是独一无二的,并提供恢复慢性乙型肝炎疲惫的T细胞,从而使恢复对乙肝病毒适应性免疫的一个极好的机会,并提供实现持续病毒学缓解一个公平的机会。使用任何方法来阻止免疫调节中心机制与没有料到的并发症。这些构成与身患绝症的癌症患者相比,可能无法接受,原本健康的慢性乙型肝炎患者的风险状况。用抗PD-1抗体的提供阻断PD-1/ L12相互作用,如PD-L1块将仍然允许PD-1-PDL2相互作用,然而,抗PD-1抗体已与最不利的相关联的最大希望事件,并且可能需要在应用之前作为治疗剂慢性乙型肝炎进一步研究。
Tregs. Tregs consist of different T-cell subpopulations, including naturally occurring CD4+ CD25+ Tregs, induced Tregs (IL-10 producing CD4+ type I Tregs (Tr1) and T helper type 3 (Th3) cells), and CD4+ CD25+ T cells that develop in the periphery by conversion of CD4+ CD25− T cells. Experimental data suggest that circulating CD+ CD25+ Tregs may suppress HBV-specific T-cell responses in CHB patients resulting in persistence of HBV.[158] 调节性T细胞。调节性T细胞包括不同的T细胞亚群,包括天然存在的CD4 + CD25 +调节性T细胞,诱导调节性T细胞(IL-10的CD4+ I型调节性T细胞(Tr1的)和T辅助细胞类型3(Th3的)细胞)和CD4 + CD25 + T细胞的发展中的周由转换的CD4 + CD25 - T细胞。实验数据表明,循环CD+ CD25 +调节性T细胞可以抑制产生HBV的持续性慢性乙型肝炎患者HBV特异性T细胞应答。[158]
Despite all the above observations, a study on WHV-infected woodchucks, treated with IL-12 in combination with a TGF-β inhibitory peptide or Treg depletion showed that TGF-β inhibition or Treg depletion had no antiviral effect, instead an enhancement of the intrahepatic tolerogenic environment was observed.[159] Since there are no distinct phenotypic characteristics of Tregs available to target, it will be a difficult task to interfere with the function of Tregs in vivo.
尽管所有的上述意见,对WHV感染的土拨鼠,与IL-12与TGF-β抑制肽或调节性T细胞耗竭结合治疗的研究表明,TGF-β抑制或调节性T细胞耗竭没有抗病毒的作用,的而不是增强肝内致耐受环境进行了观察。[159]由于存在调节性T细胞没有明显的表型特征可用于靶向,这将是一项困难的任务干扰Treg细胞在体内的功能。
Therapeutic Vaccinations. Therapeutic vaccination presents a promising strategy in approach towards HBV eradication. As discussed before, HBV-specific T-cell exhaustion due to persistent antigen stimulation is considered a major determinant of HBV persistence or chronicity. A therapeutic vaccine, which could induce a potent CD4+ T-cell response, counteract immune tolerance, activate humoral immune response and stimulate CD8+ T cells directed against one or more HBV antigens, could achieve sustained control of CHB. During the past several years, different therapeutic vaccines have been developed and investigated in CHB patients with different clinical outcomes. 治疗性疫苗。治疗性疫苗在呈现对乙肝病毒消灭的办法有前途的战略。如之前所讨论的,HBV特异性T细胞耗竭由于持续的抗原刺激被认为是乙型肝炎病毒的持久性或慢性的主要决定因素。一种治疗性疫苗,其可以诱导一种强的CD4 + T细胞应答,抵消免疫耐受,激活的体液免疫反应和刺激的CD8 + T细胞针对一种或多种HBV抗原,可以实现CHB的持续控制。在过去的几年中,不同的治疗性疫苗已经开发和研究了慢性乙型肝炎患者具有不同的临床结果。
Several categories of therapeutic vaccines are being developed for CHB infection, which includes: vaccines based on recombinant HBV proteins, HBV-envelope subviral particles, naked DNA eventually combined with viral vectors and vaccines based on T-cell peptide epitopes derived from different HBV proteins.[160–162]
正在开发用于慢性乙型肝炎感染,其中包括几种类型的治疗性疫苗:基于重组HBV蛋白疫苗,乙肝病毒包膜亚病毒颗粒,裸DNA,最终基于从不同的HBV蛋白的T细胞肽表位的病毒载体和疫苗的组合。 [160-162]
Early classical therapeutic vaccines were based on the HBsAg protein that proved to be excellent in terms of its prophylactic potential, however, use of these vaccines have failed to reach expectations in terms of therapeutic efficacy. Antiviral effect of conventional prophylactic HBsAg-based vaccines were only transient and did not result in sustained viral suppression.[163] This is probably because most patients with CHB have diminished HBV responses due to exhaustive T-cell response and do not respond to classical immunizations.
早期的经典治疗性疫苗,是根据乙肝表面抗原蛋白,其被证明是优异的预防潜在的术语,但是,使用这些疫苗都未能达到在治疗功效方面的期望。常规预防的HBsAg为基础的疫苗的抗病毒效果只有短暂的,并没有导致持续的病毒抑制[163]这可能是因为大多数慢性乙型肝炎患者已经减少由于详尽的T-细胞应答的HBV反应和不经典的免疫反应。
Therapeutic Vaccination Based on Recombinant HBV Proteins or HBV-envelope Sub-viral Particles. 治疗性疫苗接种。基于重组乙型肝炎病毒蛋白或乙型肝炎病毒包膜亚病毒颗粒。
Immunogenic – Complexes: A double-blind placebo-controlled Phase IIb trial of a therapeutic HBV vaccine based on antigen–antibody immune complexes (HBsAg with antiHBs immunoglobulin) has been conducted in 242 patients with CHB presenting some initial evidence of clinical and virological efficacy.[164] These patients received antigen–antibody immune complexes with alum as adjuvant with the aim of targeting DCs. The rationale of this combination was based on the hypothesis that immune complex-loaded DCs were superior in efficiently priming HBV-specific CD8+ cytotoxic T cells responses in vivo compared with naturally occurring immune complexes.[165] 免疫原性 - 配合:基于抗原 - 抗体免疫复合物治疗性乙肝疫苗的双盲安慰剂对照的IIb期临床试验(乙肝表面抗原与免疫球蛋白antiHBs)已在242例进行了介绍慢性乙型肝炎的临床和病毒学疗效一些初步的证据。 [164]这些患者接受抗原 - 抗体免疫复合物用明矾为佐剂并靶向DC的目的。这个组合的基本原理是基于这一假设的免疫复合物负载的DC是在有效地吸HBV特异性CD8 +细胞毒性T细胞在体内的反应与天然存在的免疫复合物进行比较优越。[165]
However, the results of Phase III clinical trial failed to show the therapeutic efficacy of immune complex-based vaccination.[166]
然而,III期临床试验的结果没有显示出的免疫复合物为基础的疫苗接种的治疗效果。[166]
HBsAg & HBcAg Combination: Another approach involves the nasal HBV vaccine candidate, comprising HBsAg and core (HBcAg) as vaccine antigens. The nasal HBV vaccine candidate is based on the results of preclinical studies that have demonstrated a good immunogenicity and safety profile. Recombinant HBcAg can act as a potent Th1 adjuvant to HBsAg, as well as a strong immunogen.[167,168] This was tested in a Phase I double-blinded, placebo-controlled randomized clinical trial in healthy volunteers that demonstrated the safety and immunologic efficacy of this combination approach.[169] At present, a Phase III clinical trial is ongoing with HBsAg/HBcAg-based combined vaccine through the nasal and subcutaneous route in CHB patients and results are awaited.[170] 的HBsAg和HBcAg的组合:另一种方法涉及到鼻乙肝疫苗候选,包括乙肝表面抗原和核心(核心抗原)作为疫苗抗原。鼻乙肝疫苗的候选是基于临床前研究已经表现出了良好的免疫原性和安全性的结果。重组核心抗原可以作为一种有效的Th1佐剂乙肝表面抗原,以及强大的免疫原。[167168]这是在I期双盲,安慰剂对照的随机临床试验,在证明的安全性和免疫效果的健康志愿者进行测试这种组合方式。[169]目前,III期临床试验正在进行用HBsAg / HBcAg的基础,通过慢性乙型肝炎患者和结果期待已久的鼻腔,皮下途径联合疫苗。[170]
Whole Recombinant Yeast-based Therapeutic Vaccine: A yeast-based immunotherapy platform, Tarmogens (targeted molecular immunogen), is currently under development. Tarmogen incorporates multiple viral antigens, expressing HBV X, surface (S), and core Ags (X-S-core) and has been shown to induce both CD4+ and CD8+ T-cell responses in healthy and CHB patients ex vivo. TheGS-4774 is a tarmogen that consists of whole, heat-killed, recombinant Saccharomyces cerevisiae yeast, which is genetically modified to express HBV antigens. Use of entire yeast results in preferential uptake/processing by dendritic cells rather than B cells to present HBV antigens to T cells, theoretically resulting in a more efficacious cellular immunity compared with a predominantly humoral immunity with the use of subunit vaccines.
整体重组酵母为基础的治疗性疫苗:酵母为基础的免疫治疗平台,Tarmogens(分子靶向免疫),目前正在开发中。 Tarmogen结合多种病毒抗原,表达HBV X,表面(S),以及核心AGS(XS芯),并已显示出诱导CD4 +和CD8 + T细胞应答在健康和慢性乙型肝炎患者体外。在GS-4774是一种tarmogen,由全,热灭活的重组酿酒酵母的酵母,其是通过基因修饰以表达HBV抗原。使用的整个酵母导致优先摄取/加工由树突状细胞而不是B细胞呈现HBV抗原给T细胞,理论上产生更有效的细胞免疫由于主要是体液免疫与使用亚单位疫苗进行比较。
Guo et al. demonstrated antigen-specific T-cell responses generated in mice immunized with two candidate vaccines mentioned above.[171] In addition, the data showed that GS 4774 (X-S core) significantly protected mice from tumors engineered to express HBV antigens. In a second study by Kemmler et al., both tarmogens elicited HBV-specific T-cell responses ex vivo in samples collected from healthy individuals and donors with CHB.[172]
Guo等。证实在免疫的上述两个候选疫苗的小鼠中产生的抗原特异性T细胞应答。[171]此外,该数据表明,GS4774(XS芯)从肿瘤显著保护小鼠改造以表达HBV抗原。在第二项研究由KEMMLER等,均tarmogens引起HBV特异性T细胞反应离体健康者和捐助者与慢性乙肝收集的样本。[172]
Recently, murine and human immunogenicity models were used to evaluate the type and magnitude of HBV-Ag specific T-cell responses elicited by the vaccine. Mice immunized with yeast expressing X-S-core showed T-cell responses to X, S and core. Both CD4+ and CD8+ T-cell responses were observed. Human T cells transduced with HBc18–27 and HBs183–91 specific T-cell receptors (TCRs) produced IFN-γ following incubation with X-S-core-pulsed DCs. Furthermore, stimulation of peripheral blood mononuclear cells isolated from CHB patients or from HBV vaccine recipients with autologous DCs pulsed with X-S-core or a related product (S-core) resulted in pronounced expansions of HBV Ag-specific T cells possessing a cytolytic phenotype. These data indicate that X-S-core-expressing yeast elicits functional adaptive immune responses. This therapeutic vaccine seems promising in inducing HBV-specific T-cell responses in patients with CHB [KING TH ET AL . UNPUBLISHED DATA].
最近,小鼠和人的免疫原性的模型,用于评估的类型和由疫苗引起的HBV-银特异性T细胞应答的量级。免疫小鼠酵母表达XS核表明T细胞反应,以X,S和核心。 CD4 +和CD8 + T细胞应答的变化。人类T细胞转导的HBc18-27和HBs183-91特异性T细胞受体(TCR)产生的IFN-γ孵育后用XS-核 - 脉冲的DC。此外,外周血单个核细胞慢性乙肝患者或乙肝疫苗接种者有脉冲与XS核或相关产品(第芯)自体DC隔离的刺激导致乙肝病毒抗原特异性T细胞具有溶细胞表型的显着扩张。这些数据表明,XS核表达酵母引发功能性免疫应答。这种治疗性疫苗似乎有希望在诱导HBV特异性T细胞应答在慢性乙型肝炎患者[KING TH等。未发表的数据。
Tarmogen, GS-4774 is being tested in combination with direct-acting antivirals in CHB patients to determine if the combination can increase rates of HBsAg seroconversion. A Phase I trial on GS-4774 has been completed and a Phase II, randomized, open-label study to evaluate the safety and efficacy of GS-4774 for the treatment of virally suppressed subjects with CHB is currently ongoing. Ongoing Phase II studies will determine whether yeast-based vaccines would result in unexpected allergic adverse reactions to common yeast infections in these patients.
Tarmogen,GS-4774正与慢性乙型肝炎患者直接作用的抗病毒药物的组合进行测试,以确定该组合可以提高HBsAg的血清转化率。对GS-4774 I期临床试验已经完成,第二阶段,随机,开放标签研究,以评估GS-4774的安全性和有效性的病毒抑制者与慢性乙肝的治疗是目前正在进行中。正在进行的II期研究中,将决定基于酵母的疫苗是否会导致意想不到的过敏性不良反应在这些患者中常见的真菌感染。
Adeno-virus-based Therapeutic Vaccination: TG1050 is a therapeutic vaccination based on a recombinant nonreplicative human adenovirus serotype 5, expressing multiple specific HBV antigens (truncated core, modified polymerase and HBsAg domains). The product has been designed to prime de novo and/or stimulate functional T cells expected to control the HBV replication and to clear HBV. In an experiment by Perrine Martin et al, TG1050 was found to induce high levels of T cells targeting core, polymerase and HBsAg domains in naive mice. In the Adeno-associated virus (AAV) tolerant mouse model, a single injection of TG1050 was shown to induce functional and long lasting T cells producing IFN-γ, TNF-α and IL-2, which were detected in spleen and liver without elevation of ALT. Experiments are ongoing to analyze immunological and virological effects of multiple injections of TG1050 as well as longer follow-ups.[173] t of CHB patients. 腺病毒为基础的治疗性疫苗:TG1050是基于一个非复制性重组人腺病毒血清型5治疗性疫苗,多表达乙肝病毒特异性抗原(截断核心,修改聚合酶和乙肝表面抗原域)。该产品已被设计成素从头和/或刺激预期来控制病毒复制和清除HBV的功能性T细胞。在实验中通过珀赖恩Martin等,TG1050,发现诱导高水平靶向芯,聚合酶和HBsAg的结构域在幼稚小鼠的T细胞。在腺相关病毒(AAV)耐受小鼠模型中,单次注射TG1050的结果显示,诱导产生IFN-γ,TNF-α和IL-2,这是在脾脏和肝脏中检测到不抬高功能和持久性T细胞的ALT键。实验正在进行中,分析免疫学和TG1050多次注射的病毒学效果以及较长的后续行动。[173] T,慢性乙肝患者 作者: kite2002005 时间: 2014-9-8 17:40
Therapeutic Vaccination Based on DNA & T-cell Peptide Epitope 治疗性疫苗基于DNA和T细胞表位肽
DNA Vaccine. DNA vaccines carry the potential to induce T-cell responses. Both CD4+ and CD8+ cells were elicited by the DNA vaccine, with the helper cells being of the Th1 phenotype, secreting IFN-γ.[174] This vaccine was shown to activate not only the T-cell responses specific to HBV but also NK cells.[175]
DNA疫苗。 DNA疫苗进行以诱导T细胞应答的潜力。 CD4 +和CD8 +细胞的DNA疫苗被引出,与辅助细胞是Th1细胞表型的,分泌的IFN-γ[174]该疫苗被证明激活不仅针对乙肝病毒的T细胞应答,而且NK细胞[175]
In one study using DNA vaccine comprising of most HBV genes encoding multiple HBV proteins (i.e., envelope, nucleocapsid and polymerase) plus genetically engineered IL-12 DNA (IL-12N222L) was used in 12 CHB carriers who were being treated with antiviral, 3TC. Detectable HBV-specific IFN-γ secreting T-cell responses were observed at the end of treatment and during a follow-up. These type 1 T-cell responses, particularly CD4 (+) memory T-cell responses were maintained for 40 weeks after the therapy was stopped and correlated with virological responses.[176]
在一项研究中使用的DNA疫苗,包括乙肝病毒大部分基因编码的多HBV蛋白(即包络,核衣壳和聚合酶)以及基因工程的IL-12的DNA(IL-12N222L)在谁正在用抗病毒药物,拉米夫定治疗慢性乙型肝炎12运营商使用。检测HBV特异性IFN-γ分泌T细胞反应均在治疗结束时和在一个后续观察。这些类型1的T细胞应答,特别是CD4 +记忆T细胞应答维持40周的治疗停止,并与病毒学应答相关联后,[176]
T-cell Peptide Epitope Vaccine. In a Phase I trial conducted in healthy volunteers, a polyepitope-based vaccine was used. This consisted of a DNA vector coding for a string of 30 HBV-derived cytotoxic T-cell epitopes linked to 16 Th epitopes, which was expected to be presented to T cells by a large number of HLA molecules. This vaccine was found to be safe and well tolerated in all healthy volunteers.[177] T细胞表位肽疫苗。在健康志愿者中进行的I期临床试验中,多表位为基础的疫苗使用。这由一个DNA载体编码为30的HBV-衍生的细胞毒性T细胞表位的字符串链接到16的Th表位,这是预期到由大量的HLA分子呈现给T细胞。这种疫苗被认为是安全且耐受良好的所有健康的志愿者。[177] 作者: StephenW 时间: 2014-9-8 17:52
Combination Approach组合方法
Chronic HBV infection is associated with T-cell exhaustion in the presence of HBV replication and antigen load. Reducing the viral load with antiviral therapy prior to vaccination could provide improvement in vaccine efficacy. Studies have shown that therapeutic vaccinations are more effective when used in patients with low HBV DNA load in serum at the start of treatment.[178,179] Recent vaccination trials are based on this approach. Therapeutic vaccination is used concomitantly with antiviral agents to induce T-cell restoration with suppression of viral replication and antigen load, which is an important factor responsible for induction of tolerance.[180,181]
慢性HBV感染与HBV复制和抗原负载的存在的T细胞衰竭有关。降低病毒载量与抗病毒治疗前,接种疫苗能提供改善疫苗的有效性。研究表明,在低的患者的HBV DNA载在血清在治疗开始时使用治疗性的疫苗接种是更有效的。[178179]最近接种试验是基于这样的方法。治疗性疫苗接种的同时使用与抗病毒剂,以诱导T细胞的恢复和抑制病毒复制和抗原负荷,这是负责诱导耐受性的一个重要因素。[180181] 作者: 秋路 时间: 2014-9-8 17:54
Since sustained virologic remission of HBV does not occur as a result of existing treatment, novel strategies will have to be adopted. A learned approach to achieve this goal is to circumvent the various factors by which HBV establishes chronic infection in humans. Targeting a combination of viral and host factor offers the best possible chance for accomplishing this objective. Safety of a variety of these approaches, especially those targeting host immune responses have not yet been established. If these approaches are safe, they offer a unique perspective to target host immunity against HBV proteins and eventually develop protective immunity and control of HBV replication. A key viral target remains cccDNA, which requires unique approaches, which have shown in vitro effectiveness. The challenges remain on developing successful delivery system that allows every single infected cell to be subjected to disruption of ccDNA. In reality, a combination approach would be the most appropriate, effective and pragmatic technique to achieve sustained virologic remission of most CHB patients (Figure 1 & Table 1 & Table 2).
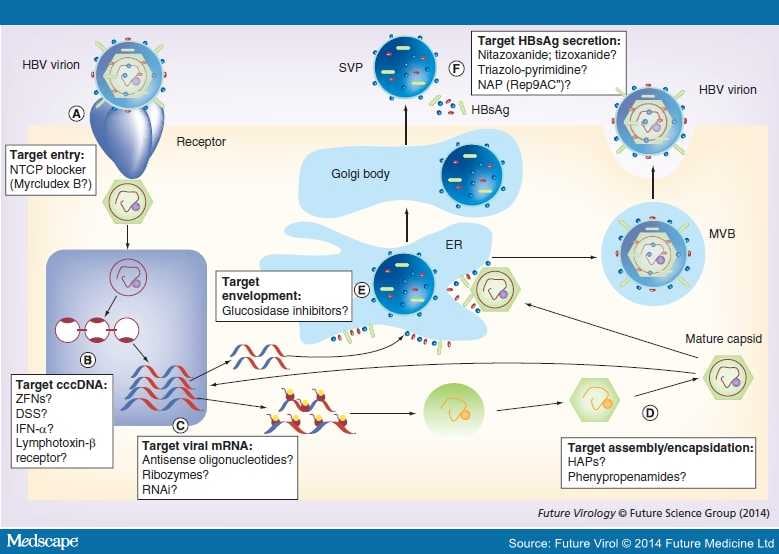
HBV lifecycle showing novel approaches for viral targets. (A) Viral entry; (B) cccDNA; (C) viral mRNA; (D) assembly/encapsidation; (E) envelopment in ER; (F) SVP and HBsAg. Mature viral capsid enveloped in ER and is secreted through MVBs. SVPs are processed and secreted through the ER/Golgi pathway. Possible novel targets are represented by question marks.
长期随访研究显示,在 HBeAg 阳性、使用 PEG-IFN 伴或不伴 3TC 治疗获得持久病毒抑制的长期临床缓解患者中,其 HBsAg 血清转换率增高、肝组织学改善。基因型 A 或 B 的应答率也高于 C 或 D 型,且 A 和 B 型患者基线 HBV DNA 水平较低但 ALT 值较高。PEG-IFN 因其耐受性差和不良事件多而受到使用限制。
病毒核心 mRNA 编码核心蛋白(主要的核衣壳蛋白)和 DNA 聚合酶(用于逆转录 RNA 前基因组),并充当前基因组 RNA,后者为逆转录的模板。前核心 mRNA 编码前核心蛋白(在内质网中处理产生 HBeAg)、基础核心基因启动子(BCP,核苷酸 1744-1804,位于 X ORF,控制前 C 区和 C 区的转录活动)。目前已报道有多中前核心和核心区突变。
基因型 A 是急性 HBV 感染后进展至慢性感染且持续存在的独立风险因子。基因型为 A、D 的急性 HBV 感染发展至慢性感染的机率高于 B 和 C 型。基因型 C 和 D 的患者自发性 HBeAg 血清转换率低于 A 型和 B 型。HBV 基因型与前 C 和 BCP 突变有明确的关系。基因型 C 有更高的频率出现 BCP A1762T/G1764A 双突变和前 -S 缺失,同时其病毒载量也高于基因型 B。
类似的,基因型 D 出现 BCP A1762T/G1764A 双突变的频率高于基因型 A。基因型 C 和 D 与严重的肝脏疾病如肝硬化和 HCC 有关。基因型 A 和 B 对干扰素治疗的应答反应优于 C 和 D 型,但对 NAs 的应答无差异。
目前已明确这种基因型通过允许更多的蛋白泛宿主结合至此等位基因来影响宿主反应,导致在带有此有利等位基因的患者上有更广泛的 T 细胞反应,从而使感染自限化。另一研究 CHB 患者同胞的全基因关联研究发现,21 号染色体上横跨 IFN-α 受体 II 和 IL-10 受体 II 的单个核苷酸多态性的连锁区与慢性感染相关。
d、调节性 T 细胞。调节性 T 细胞(Treg)由不同的 T 细胞亚群组成,包括自然发生 CD4+ CD25+ Tregs,诱导型 Tregs(产 IL-10 CD4+ I 型 Tregs(Tr1)和 T 辅助 III 型细胞),以及从 CD4+ CD25- 转换而来在外周血中发育的 CD4+CD25+T 细胞。实验性数据表明,循环 CD4+CD25+Tregs 或许能抑制 CHB 患者中的 HBV 特异性 T 细胞应答,从而导致 HBV 持续存在。
GS-4774 为一种 tarmogen,由整个热灭活重组酵母属啤酒酵母菌组成,而后者可进行基因修饰从而表达 HBV 抗原。使用整个酵母菌可使病原体更好地被树突状细胞摄取或处理,以将 HBV 抗原呈递给 T 细胞,与使用亚单位疫苗所致的主导性体液免疫相比,理论上其可导致更有效的细胞免疫。
Guo 等发现,使用上述备选疫苗免疫的小鼠可产生抗原特异性 T 细胞反应。此外,数据提示,GS 4774 (X-S core) 可明显地保护小鼠免受表达 HBV 抗原的肿瘤侵袭。Kemmler 等人所进行的第二项研究发现, tarmogens 可在体外诱导样本产生 HBV 特异性 T 细胞反应,这些样本是从健康人和 CHB 患者收集而来的。
最近,鼠类和人类免疫原性模型被用来评估由疫苗所诱导产生的 HBV 抗原特异性 T 细胞应答的类型和强度。使用 表达 X-S-core 的酵母菌免疫的小鼠中发现了针对 X、S 和 core 的 T 细胞应答。CD4+ 和 CD8+T 细胞反应均可观察到。转导了 HBc18–27 和 HBs183–91 特异性 T 细胞受体的人 T 细胞在添加 X-S-core DCs 孵育后产生了 IFN-γ。
而且,从 CHB 患者中分离所得的外周血单个核细胞,或使用添加了 X-S-core 或相关产物(S-core)的同源性 DCs 进行 HBV 疫苗接种者中分离的外周血单个核细胞产生了显著的 HBV 抗原特异性 T 细胞扩增,这些 T 细胞拥有细胞溶解表型。这些数据提示,表达 X-S-core 的酵母菌产生功能性适应性免疫反应。这些治疗性疫苗在诱导 CHB 患者产生 HBV 特异性 T 细胞应答上似乎很有前景。
Tarmogen GS-4774 正在 CHB 患者身上与直接抗病毒药物联合使用,以明确联合用药是否能提高 HBsAg 的血清转换率。一个关于 GS-4774 的 I 期临床试验已经结束,评估 GS-4774 治疗病毒抑制状态 CHB 患者的疗效和安全性的 II 期随机、开放标签试验正在进行中。正在进行的 II 期试验将会明确以酵母菌为基础的疫苗是否会在这些患者感染普通酵母菌感染时导致不可预测的过敏不良反应。
基于腺病毒的治疗性疫苗。TG1050 是一种基于重组非复制性人腺病毒血清型 5、表达多种特异性 HBV 抗原(截短型核心抗原、修饰后的多聚酶和 HBsAg 区抗原)的治疗性疫苗。这些产物已经被设计来诱导 T 细胞重新合成或刺激能控制 HBV 复制和清除 HBV 的功能性 T 细胞增殖。
Perrine Martin 等进行的一个试验发现 TG1050 能在初次接受试验的小鼠中诱导高水平的针对核心、多聚酶和 HBsAg 区的 T 细胞。腺相关病毒耐受的小鼠模型中,仅注射一次 TG1050 能诱导产 IFN-γ,、TNF-α 和 IL-2 的功能性和持久性 T 细胞,后者可在脾和肝脏中发现而 ALT 正常。分析多剂量注射 TG1050 的免疫学和病毒学效应的研究正在进行。
b、基于 DNA 和 T 细胞肽表位的治疗性疫苗
DNA 疫苗。DNA 疫苗具有诱导 T 细胞应答的潜能。注射 DNA 疫苗可产生 CD4+ 和 CD8+ 细胞,以及带有 Th1 表型、分泌 IFN-γ的辅助细胞。DNA 疫苗不仅能激活 HBV 特异性 T 细胞应答,也能激活 NK 细胞。
一项使用包含有编码多种 HBV 蛋白的大部分 HBV 基因的 DNA 疫苗联合基因构建的 IL-12 DNA 的研究在 12 名使用 3T 抗病毒治疗的 CHB 携带者中进行。治疗结束和随访时在患者体内发现了可检测的 HBV 特异性的分泌 IFN-γ的 T 细胞反应。这些 I 型 T 细胞反应,特别是 CD4+ 记忆 T 细胞应答在治疗结束后仍可维持 40 天,并与病毒学应答相关。
T 细胞肽表位疫苗。在一个 I 期临床试验中,对健康志愿者注射了多表位疫苗,后者由编码一系列的与 16Th 表位相关的来源于 30 种 HBV 细胞毒性 T 细胞表位的 DNA 载体组成,其有望通过大量的 HLA 分子递呈抗原至 T 细胞。在健康志愿者中这种疫苗安全、耐受性良好。
c、联合治疗方法
慢性 HBV 感染与 HBV 复制存在和抗原负载所致的 T 细胞耗竭相关。在注射疫苗前使用抗病毒治疗降低病毒载量能提高疫苗疗效。研究表明,治疗性疫苗在治疗开始时使用到低 HBV DNA 病毒载量的患者上时更有效。新近的试验都是基于此方法。治疗性疫苗与抗病毒药物一起使用可诱导病毒复制和抗原负载受到抑制下的 T 细胞恢复,后者在耐受性的诱导中起重要作用。